Introdução
Aditivos alimentares que interagem com a resposta imune têm considerável potencial para reduzir a susceptibilidade das aves a doenças infecciosas. Neste contexto tem-se estudado a relação entre sistema imune e saúde animal frente ao uso de prebióticos como os mananoligossacarídeos (MOS). Estes compostos são derivados da parede celular de leveduras Saccharomyces cerevisiae que, quando adicionado a ração animal, afetam beneficamente o hospedeiro por estimular o crescimento seletivo e/ou à atividade de um número limitado de bactérias da microbiota intestinal (Gibson & Roberfroid, 1995). O estudo de seu modo de ação e demais benefícios é de particular interesse para a avicultura, uma vez que os MOS não só alteram a microbiota intestinal (Fairchild et al., 2001), mas também apresentam propriedades imunomoduladoras (Savage et al., 1996; Shashidhara & Devegowda, 2003). Sendo assim, o objetivo deste estudo foi quantificar os células caliciformes e linfócitos T na mucosa intestinal de frangos de corte suplementados com MOS, acompanhado de vacina e desafio com Salmonella Enteritidis (SE).
Materiais & Métodos
Quarenta e cinco frangos de corte Cobb 500 de um dia de idade foram distribuídos em 3 tratamentos com 15 repetições cada. As aves, identificadas por anilhas numeradas, foram pesadas semanalmente e vacinadas aos 14 e 28 dias com vacina comercial inativada contraSE.Aos 42 dias as aves foram inoculadas com 1 mL de solução contendo 105 UFC de SE por via oroesofágica. As dietas utilizadas foram: Controle: ração controle a base de milho e farelo de soja, MOS++, ração controle com maior nível de MOS (2kg/Ton 1-21dias e 1kg/Ton 22-56 dias) e MOS+, ração controle com menor nível de MOS (1kg/Ton 1-21dias e 0,5 kg/Ton 22-56 dias). Aos 7, 37 e 56 dias cinco aves por tratamento foram necropsiadas para colheita de fragmentos de ceco. Foram realizadas análises histológicas para a quantificação de células caliciformes e análise imunohistoquimica para a quantificação de linfócitos T CD3. Os resultados foram analisados por ANOVA (P<0.05) e as médias diferentes submetidas ao teste Exato de Fischer.
Resultados & Discussão
Na Tabela 1 são apresentados os valores de ganho de peso nos períodos de 1 a 21 dias, 1 a 42 dias, 1 a 49 dias e 1 a 56 dias de vida das aves. Nos períodos de 1 a 21 dias e 1 a 42 dias de idade, o grupo MOS ++ apresentou pior ganho de peso quando comparado aos outros grupos experimentais, porém não foi observada diferença estatística em outros períodos para análise de ganho de peso.
Tabela 1. Ganho de peso das aves nos diferentes períodos de crescimento das aves nos grupos experimentais
Média ± desvio. a, b Letras diferentes na mesma coluna indicam diferença significativa (P< 0,05).
Na Tabela 2 são apresentados os resultados das análises microbiológicas para contagem de SE. Não houve diferença estatística entre os grupos experimentais nas coletas realizadas aos 2 e 7 dias após desafio com SE, porém o grupo MOS+ reduziu a excreção fecal de SE aos 14 dias após o desafio quando comparado aos outros grupos experimentais.
Tabela 2. Contagem de Salmonella Enteritidis, Log 10 UFC, em suabes de cloaca coletados aos 2, 7 e 14 dias após desafio (AD) nos diferentes grupos experimentais
a,b Letras diferentes na mesma coluna indicam diferença significativa (P< 0,05).
Tabela 3. Contagem de células CD3+ no íleo e cecos das aves aos 7, 37 e 56 dias de vida nos diferentes grupos experimentais (Imunohistoquimica, 100X)
Média ± desvio. a, bLetras diferentes na mesma coluna indicam diferença significativa (P< 0,05).
No presente estudo, a análise dos resultados de ganho de peso de 1 a 21 e 1 a 42 dias, em comparação com a quantificação de células CD3+ fornece uma relação direta entre estes dois parâmetros, onde a redução no ganho de peso ocorre simultaneamente ao aumento na quantificação de células CD3+ no grupo MOS++ (Tabela 1 e Tabela 3). Sugere-se então que houve uma interferência do processo de vacinação nestes resultados de ganho de peso, pois as doses vacinais foram aplicadas aos 14 e 28 dias de vida das aves. Segundo Klassing (1998) a resposta imune pode aumentar a necessidade de recursos orgânicos pelo animal e assim, afetar o desempenho das aves, como também sugerido por Dibner et al. (1998) e Kogut & Klasing (2009).
O sistema imune inato das aves é capaz de reconhecer estruturas características de microorganismos patogênicos e que não estão presentes nas células de aves e mamíferos como lipopolissacarídeos e mananoligossacarideos (Abbas, 2000). A ação protetora dos derivados de parede celular de levedura tem sido descrita como imunomodulação não específica devido ao envolvimento de diferentes vias, incluindo a ativação de macrófagos, estimulação de células-T, estimulação do sistema reticulo endotelial, ativação de células NK, ativação de vias complementares clássicas e alternativas (Zekovic et al., 2005) e aumento da produção de anticorpos (Shashidhara & Devegowda, 2003).
A Figura 1 apresenta os valores da contagem de células CD3+ e da quantificação de células caliciformes por vilo no ceco das aves. Nesta podemos observar uma relação inversa entre o número de células caliciformes e células CD3+ na mucosa intestinal das aves, onde após a aplicação das doses vacinais houve um aumento do número de células CD3+ e redução do número de células caliciformes. Após 14 dias do desafio com SE (56 dias) a mucosa intestinal das aves apresentou aumento do número de células caliciformes e redução de células CD3+ quando comparado aos valores observados aos 37 dias.
Figura 1. Quantificação de células CD3+ e células caliciformes nos cecos das aves aos 7, 37 e 56 dias de vida nos diferentes grupos experimentais.
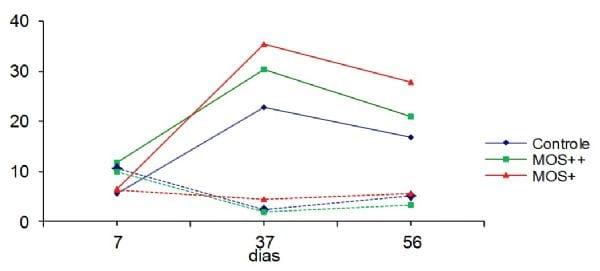
Nos fragmentos de ceco foi observado, no grupo controle e MOS++, redução no número de células caliciformes por vilo aos 37 dias e aumento deste número aos 56 dias ou 44 dias após desafio com SE. O grupo MOS+ não apresenta esta tendência, visto que estas aves quase sempre apresentaram maior número de células caliciformes quando comparadas aos grupos controle e MOS++.
Com base nos resultados obtidos na quantificação de células CD3+ do íleo, sugere-se que maior quantidade de MOS na ração provoca uma imunoestimulação, enquanto que a menor quantidade suplementada via ração provoca uma imunomodulação. Observou-se que existe uma relação inversa entre o número de células CD3+ e a expressão de células caliciformes na mucosa intestinal, uma vez que houve aumento de células CD3+ e redução do número de células caliciformes após a vacinação. Nas aves do grupo suplementado com menor nível de mananoligossacarideos (MOS+) isto também ocorre, mas com pequena variabilidade, a qual pode ser benéfica para o desempenho animal. A camada de muco, produzida pelas células caliciformes, atua como um meio de proteção físico e biológico e também é um componente da resposta imune inata que é regulada em resposta à inflamação e infecção (Uni et al., 2003).
Os resultados referentes aos grupos suplementados sugerem que um maior nível de MOS relaciona-se mais claramente com os processos fisiológicos da mucosa intestinal de frangos de corte, enquanto que menores níveis de MOS parecem ser mais eficientes no controle da excreção de SE.
A suplementação com MOS desencadeou um efeito dose-resposta na resposta imunológica das aves, pois houve uma maior contagem de linfócitos T no íleo do grupo MOS ++.
Conclusões
A suplementação com mananoligossacarideos (1kg/Ton 1-21 dias e 0,5kg/Ton 22-56dias) na ração das aves foi capaz de diminuir a excreção fecal de SE, 14 dias após o desafio, quando comparadas ao grupo controle. Os resultados sugerem uma relação inversa entre a presença de células CD3+ e a expressão de células caliciformes ou imunidade inata.
Bibliografia
Abbas AK. 2000. Innate Immunity. pp. 270-290. In: Cellular and molecular immunology, Abbas AK (ed.).Saunders WB, Philadelphia.
Dibner JJ, Knight CD, Kitchel ML, Atwell CA, Downs AC, Ivey FJ. 1998. Early feeding and development of the immune system in neonatal poultry. J Appli Poult Res. 7:425-436.
Fairchild AS, Grimes JL, Jones FT, Wineland MJ, Edens FW, Sefton, AE. 2001. Effects of hen age, Bio-Mos, and Flavomycin on poultry susceptibility to oral Escherichia coli challenge. Poult Sci. 80:562-571.
Gibson GR & Roberfroid MB. 1995. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J Nutr. 125:1401-1012.
Klasing KC. 1998. Nutritional modulation of resistance to infectious diseases. Poult Sci 77:1119-1125.
Kogut MH & Klasing K. 2009. An immunologist's perspective on nutrition, immunity, and infectious diseases: Introduction and overview. J Appl Poult Res 18:103-110.
Savage TF, Cotter PF, Zakrzewska EI. 1996. The effect of feeding mannanoligosaccharide on immunoglobulins, plasma IgG and bile IgA of wrolstad MW male turkeys. Poult Sci 75(Suppl. 1):143.
Shashidhara RG & Dewegowda G. 2003. Effect of dietary mannan oligosaccharide on broiler breeder production traits and immunity. Poult Sci. 82:1319-1325.
Uni Z, Smirnov A, Sklan D. 2003. Pre- and Posthatch development of goblet cells in the broiler small intestine: Effect of delayed access to feed. Poult Sci. 82:320-327.
Zekovic DB, Kwiatkowski S, Vrvic MM, Jakovljevic D, Moran CA. 2005. Natural and modified (1/3)-β -D-Glucans in health promotion and disease alleviation. Crit Rev Biotechnol 25:205-230.






