Aminoácidos limitantes na nutrição de suínos
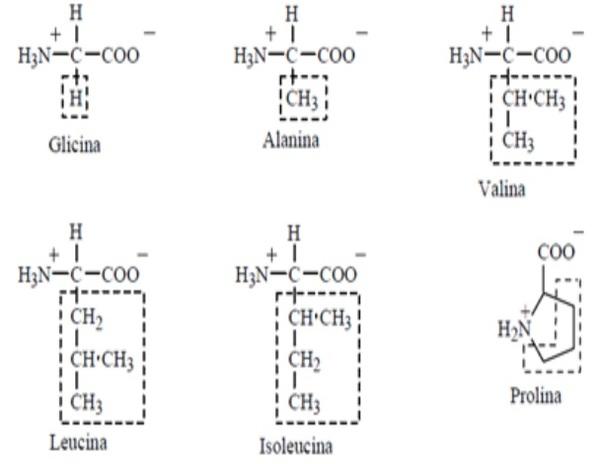
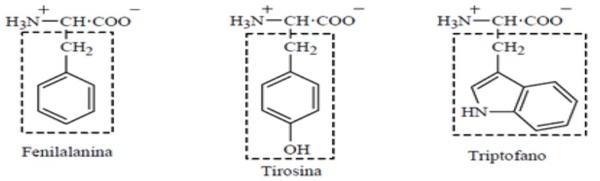
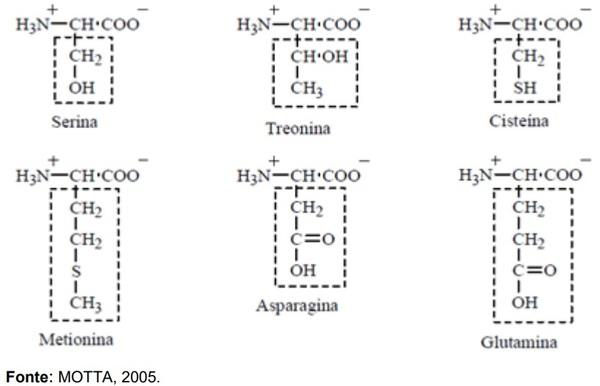

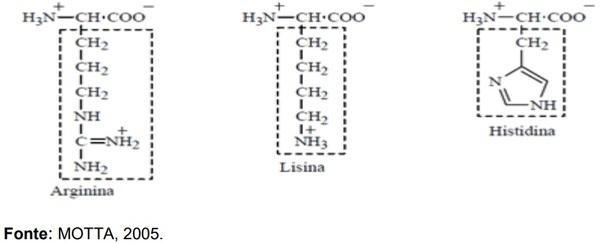
ADEOLA, O.; LAWRENCE, B. V.; CLINE, T. R.
Availability of amino acids for 10-to 20-kilogram pigs: lysine and threonine in soybean meal. Journal of animal science, v. 72, n. 8, p.
2061-2067, 1994.
AJINOMOTO – Boletim Técnico 10. Exigências de treonina para suínos. Benefícios da suplementação de L-treonina. 2003. Acessado em 29/04/2015. Online. Disponível em: www.lisina.com.br.
ALMEIDA SANTOS, F.; DONZELE, J. L.; DE
OLIVEIRA, R. F. M.; DE OLIVEIRA SILVA, F. C.;
DE ABREU, M. L. T.; SARAIVA, A. Níveis de treonina digestível em rações para suínos machos castrados de alto potencial genético na fase dos 95 aos 125 kg. Revista Brasileira de
Zootecnia, v. 39, n. 3, p. 526-531, 2010.
ALMEIDA SANTOS, F.; DONZELE, J. L.; DE
OLIVEIRA, R. F. M.; DE OLIVEIRA SILVA, F. C.;
DE ABREU, M. L. T.; SARAIVA, A. Níveis de lisina digestível para suínos machos castrados de alto potencial genético dos 95 aos 125 kg.
Revista Brasileira de Zootecnia, v. 40, n. 5, p.
1038-1044, 2011a.
ALMEIDA SANTOS, F.; DONZELE, J. L.; DE
OLIVEIRA, R. F. M.; DE OLIVEIRA SILVA, F. C.;
DE ABREU, M. L. T.; SARAIVA, A. Levels of digestible methionine+ cystine in diets for high genetic potential barrows from 95 to 125 kg.
Revista Brasileira de Zootecnia, v. 40, n. 3, p.
581-586, 2011b.
ARAÚJO, W. A. G.; SOBREIRA, G. F. PROTEÍNA
IDEAL COMO ESTRATÉGIA NUTRICIONAL NA
ALIMENTAÇÃO DE SUÍNOS. Revista Eletrônica
Nutritime, v. 5, n. 2, p. 537-545, 2008.
BAKER, D.H. Partitioning of nutrients for growth and other metabolic functions. Poultry Science,v.70, p. 1797-1805, 1991.
BAREA, R.; BROSSARD, L.; FLOC’H, L.; PRIMOT,
Y.; MELCHIOR, D.; VAN MILGEN, J. The standardized ileal digestible valine-to-lysine requirement ratio is at least seventy percent in postweaned piglets. Journal of animal science, v. 87, n. 3, p. 935-947, 2009a.
BAREA, R.; BROSSARD, L.; FLOC’H, L.; PRIMOT,
Y.; VAN MILGEN, J. The standardized ileal digestible isoleucine-to-lysine requirement ratio may be less than fifty percent in eleven-to twentythree-kilogram piglets. Journal of animal science, v. 87, n. 12, p. 4022-4031, 2009b.
BATISTA, R.M.; DE OLIVEIRA, R.F.M.; DONZELE,
J.L.; DE OLIVEIRA, W.P.; LIMA, A.L.; DE
ABREU, M.L.T. Lisina digestível para suínos machos castrados de alta deposição de carne submetidos a estresse por calor dos 30 aos 60kg.
Revista Brasileira de Zootecnia, v.40, n.9, p.1925-1932, 2011.
BATTERHAM, E. S.; ANDERSEN, L. M.; BAIGENT,
D. R.; WHITE, E. Utilization of ileal digestible amino acids by growing pigs: effect of dietary lysine concentration on efficiency of lysine retention. British Journal of Nutrition, v. 64, p.
81, 1990.
BERTECHINI, A.G. Nutrição de monogástricos. 2ª
Edição Lavras: Editora UFLA, 2012. 373 p.
BERTO, A. D.; WECHSLER, S. F.; NORONHA, C. C.
Exigências de treonina de leitões dos 7 aos 12 e dos 12 aos 23 kg. Revista Brasileira de
Zootecnia, v. 31, p. 1176-1183, 2002.
BIKKER, P.; DIRKZWAGER, A.; FLEDDERUS, J.;
TREVISI, P.; LE HUEROU-LURON, I.; LALLES,
J. P.; AWATI, A. The effect of dietary protein and fermentable carbohydrates levels on growth performance and intestinal characteristics in newly weaned piglets. Journal of Animal
Science, v. 84, n. 12, p. 3337-3345, 2006.
BRUMANO, G.; GATTÁS, G. Fatores que inflenciam na exigência de lisina para suínos. Revista
Eletrônica Nutritime, v. 6, n. 3, p. 918-940,
2009.
CASTILHA, L. D.; POZZA, P. C.; NUNES, R. V.;
LAZZERI, D. B.; SOMENSI, M. L.; POZZA, M. S.
D. S. Levels of digestible isoleucine on performance, carcass traits and organs weight of gilts (15-30 KG). Ciência e Agrotecnologia, v.
36, n. 4, p. 446-453, 2012.
COLONI, R. D. O Papel da Fitase na Alimentação de Suínos. 2013. Coordenada por Revista
Agropecuária. Disponível em: <http://www.revistaagropecuaria.com.br/2011/12/15/o-papelda-fitase-na-alimentacao-de-suinos/>. Acesso em: 20 abr. 2015.
CORASSA, A.; KIEFER, C.; GONÇALVES, L. M. P.
Planos nutricionais de lisina para suínos da fase inicial a terminação. Archivos de zootecnia, v.
62, n. 240, p. 533-542, 2013.
DEAN, D. W.; SOUTHERN, L. L.; KERR, B. J.;
BIDNER, T. D. Isoleucine requirement of 80-to
120-kilogram barrows fed corn-soybean meal or corn-blood cell diets. Journal of animal science, v. 83, n. 11, p. 2543-2553, 2005.
DUARTE, K. F. Critérios de avaliação das exigências em treonina, triptofano, valina e isoleucina para frangos de corte de 22 a 42 dias de idade. 2009. 138 f. Tese (Doutorado) -
UNESP, Jaboticabal, 2009.
FIGUEROA, J. L.; LEWIS, A. J.; MILLER, P. S.;
FISCHER, R. L.; GÓMEZ, R. S.; DIEDRICHSEN,
R. M. Nitrogen metabolism and growth performance of gilts fed standard corn-soybean meal diets or low-crude protein, amino acidsupplemented diets. Journal of Animal Science, v. 80, n. 11, p. 2911-2919, 2002.
FIGUEROA, J. L.; LEWIS, A. J.; MILLER, P. S.;
FISCHER, R. L.; DIEDRICHSEN, R. M. Growth, carcass traits, and plasma amino acid concentrations of gilts fed low-protein diets supplemented with amino acids including histidine, isoleucine, and valine. Journal of animal science, v. 81, n. 6, p. 1529-1537, 2003.
FORTES, E. I.; DONZELE, J. L.; OLIVEIRA, R. F.
M.; SARAIVA, A.; SILVA, F. C. D. O.; SOUZA, M.
F. D. Sequências de lisina digestível para suínos de duas linhagens selecionadas para alta deposição de carne. Revista Brasileira Saúde e
Produção Animal, v. 13, p. 480-490, 2012.
GAINES, A. M.; BOYD, R. D.; JOHNSTON, M. E.;
USRY, J. L.; TOUCHETTE, K. J.; ALLEE, G. L.
The dietary valine requirement for prolific lactating sows does not exceed the National Research
Council estimate. Journal of animal science, v.
84, n. 6, p. 1415-1421, 2006.
GAINES, A. M.; KENDALL, D. C.; ALLEE, G. L.;
USRY, J. L.; KERR, B. J. Estimation of the standardized ileal digestible valine-to-lysine ratio in 13-to 32-kilogram pigs. Journal of animal science, v. 89, n. 3, p. 736-742, 2011.
GATTÁS, G.; SILVA, F. C. O.; BARBOSA, F. F.;
DONZELE, J. L.; FERREIRA, A. S.; OLIVEIRA, R.
F. M. Níveis de lisina digestível em dietas para suínos machos castrados dos 60 aos 100 dias de idade. Revista Brasileira de Zootecnia, v. 41, n.
1, p. 91-97, 2012.
GIROTTO JÚNIOR, C. J.; BRUSTOLINI, P. C.;
SILVA, F. C. O.; DONZELE, J. L.; FERREIRA, A.
S.; NALON, P. M.; MOUTINHO, J. V.
Suplementação de aminoácidos para redução da proteína bruta em dietas para leitões desmamados aos 21 dias de idade. Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, v. 65, n. 4, p.1131-1138, 9 jan. 2013.
HAESE, D.; DONZELE, J. L.; DE OLIVEIRA, R. F.
M.; LOBÃO, M.; DE ABREU, T.; DE OLIVEIRA
SILVA, F. C.; SARAIVA, A. Níveis de triptofano digestível em rações para suínos machos castrados de alto potencial genético para deposição de carne na carcaça dos 60 aos 95 kg.
R. Bras. Zootec, v. 35, n. 6, p. 2309-2313, 2006.
HENRY, Y.; SEVE, B.; COLLÉAUX, Y.; GANIER, P.;
SALIGAUT, C.; JÉGO, P. Interactive effects of dietary levels of tryptophan and protein on voluntary feed intake an growth performance in pigs, in relation to plasma free amino acids and hypothalamic serotonin. Journal of Animal
Science, v.70, p.1873-1887, 1992.
HEO, J. M.; KIM, J. C.; HANSEN, C. F.; MULLAN, B.
P.; HAMPSON, D. J.; PLUSKE, J. R. Effects of feeding low protein diets on plasma urea nitrogen, faecal ammonia nitrogen, the incidence of diarrhoea and performance after weaning.
Archives of Animal Nutrition, v. 62, p. 343–358,
2008.
HTOO, J., WILTAFSKY, M. K. Roles, metabolism y antagonismos de aminoacidos de cadena ramificada en la nutrición animal. AminoNews, v.
16, n. 1, p. 25-32, 2011.
KERR, B.J.; EASTER, R.A. Effect feeding reduced protein aminoacid-supplemented diets on nitrogen and energy balance in grower pigs. Journal of
Animal Science , v.73, n.10,p.3000-3008, 1995.
KERR, B. J.; KIDD, M. T.; CUARON, J. A.;
BRYANT, K. L.; PARR, T. M.; MAXWELL, C. V.;
CAMPBELL, J. M. Isoleucine requirements and ratios in starting (7 to 11 kg) pigs. Journal of animal science, v. 82, n. 8, p. 2333-2342, 2004.
KIEFER, C.; FERREIRA, A. S.; DONZELE, J. L.;
OLIVEIRA, R. F. M. D.; SILVA, F. C. D. O.;
BRUSTOLINI, P. C. Exigência de metionina + cistina digestíveis para suínos machos castrados mantidos em ambiente termoneutro dos 30 aos
60 kg. Revista Brasileira de Zootecnia, v 34, n.
3, p. 847-854, 2005.
KIEFER, C.; FERREIRA, A. S.; OLIVEIRA, R. F. M.
D.; DONZELE, J. L.; CARRIJO, A. S.; SILVA, F.
C. D. O. Níveis de treonina digestível em dietas para fêmeas suínas de alto potencial genético em lactação sob condições de alta temperatura ambiente. Revista Brasileira de Zootecnia, v.
36, n. 05, p. 1331-1339, 2007.
KIM, B. G.; LINDEMANN, M. D.; RADEMACHER,
M.; BRENNAN, J. J.; CROMWELL, G. L. Efficacy of-methionine hydroxy analog free acid andmethionine as methionine sources for pigs.
Journal of animal science, v. 84, n. 1, p. 104-
111, 2006.
KOVAR, J.L.; LEWIS, A. J.; RADKE, T. R.; MILLER,
P. S. Bioavailability of threonine in soybean meal for young pigs. Journal of Animal Science, v.71,p.2133-2139, 1993.
LAZZERI, D. B. Níveis de isoleucina digestível para suínos machos castrados dos 15 aos 30 kg. 2011. 64 f. Dissertação (Mestrado) - Curso de
Zootecnia, Universidade do Oeste do Paraná,
Marechal Cândido Rondon, 2011.
LE BELLEGO, L.; NOBLET, J. Performance and utilization of dietary energy and amino acids in piglets fed low protein diets. Livestock
Production Science, v. 7, p. 45-58, 2002.
LEWIS, A. J. Amino acids in swine nutrition. Swine nutrition, v. 2, p. 131-141, 1991.
LI, Y. Z.; KERR, B. J.; KIDD, M. T.; GONYOU, H. W.
Use of supplementary tryptophan to modify the behavior of pigs. Journal of animal science, v.
84, n. 1, p. 212-220, 2006.
LITVAK, N.; RAKHSHANDEH, A.; HTOO, J. K.; DE
LANGE, C. F. M. Immune system stimulation increases the optimal dietary methionine to methionine plus cysteine ratio in growing pigs.
Journal of animal science, v. 91, n. 9, p. 4188-
4196, 2013.
LOHMANN, A. C.; POZZA, P. C.; POZZA, M. D. S.;
NUNES, R. V.; CASTILHA, L. D.; POSSAMAI, M.;
LAZZERI, D. B. Níveis de valina digestível para suínos machos castrados dos 15 aos 30 kg.
Archivos de zootecnia, v. 61, n. 234, p. 267-
278, 2012.
MORETTO, V.; DONZELE, J. L.; OLIVEIRA, R. F. M.
D.; FONTES, D. D. O. Níveis dietéticos de lisina para suínos da raça Landrace dos 15 aos 30 kg.
Revista Brasileira de Zootecnia, 29(3):803-809,
2000.
MOTTA, V. T. Bioquímica Básica. Caxias do Sul:
Universidade de Caxias do Sul, 2005. 332 p.
MOTTA, V. T. Bioquímica Básica. Caxias do Sul:
Universidade de Caxias do Sul, 2005. 332 p.LEAL,
E.S. Extração, obtenção e caracterização parcial de ácido fítico do germe grosso de milho e aplicação como antioxidante. 2000. Dissertação (Mestrado em Ciência de Alimentos)– Universidade
Estadual de Londrina, Londrina, 2000.
MURAKAMI, A.E. Nutrição e alimentação de codornas japonesas em postura. In: REUNIÃO
ANUAL DA SOCIEDADE BRASILEIRA DE
ZOOTECNIA, 39, 2002. Recife, Anais... Recife:
SBZ, 2002.
NATIONAL RESEARCH COUNCIL. Nutrient
Requirements of Swine. 11.ed. Washington:
National Academy, 2012. 400p.
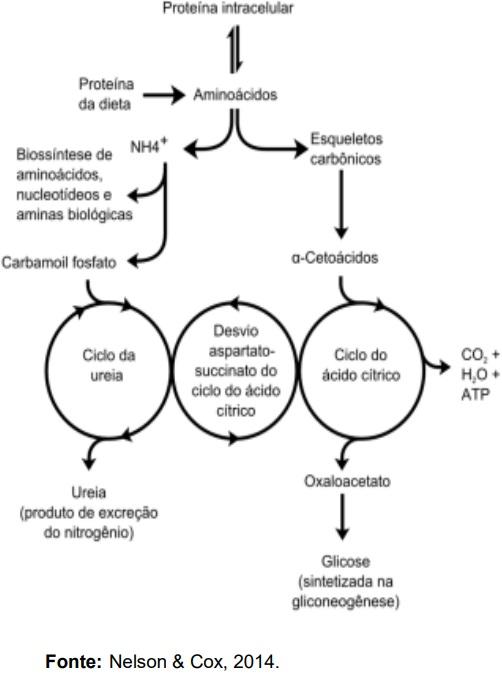
NELSON, D.L.; COX, M.M. Princípios de
Bioquímica: Lehninger. Omega, 2014.
NEME, R.; ALBINO, L. F. T.; ROSTAGNO, H. S.;
RODRIGUEIRO, R. J. B.; TOLEDO, R. S.
Determinação da Biodisponibilidade da Lisina
Sulfato e Lisina HCl com Frangos de Corte.
Revista Brasileira de Zootecnia, v. 30, n. 5, p.
1750-1759, 2001.
NYACHOTI, C. M.; OMOGBENIGUN, F. O.;
RADEMACHER, M.; BLANK, G. Performance responses and indicators of gastrintestinal health in early-weaned pigs fed low-protein amino acidsupplemented diets. Journal of Animal Science, v. 84, n. 1, p. 125-134, 2006.
OLIVEIRA, A. L. S. D.; DONZELE, J. L.; SILVA, F. C.
D. O.; OLIVEIRA, R. F. M. D.; ABREU, M. L. T.
D.; PEREIRA, A. A.; SCOTTÁ, B. A. Lisina digestível em dietas para suínos machos castrados de alto potencial Genético para deposição de carne magra na carcaça dos 60 aos
95 kg. Revista Brasileira de Saúde e Produção
Animal, v. 15, n. 4, 2014.
OLIVEIRA, A. L. S. D.; DONZELE, J. L.; OLIVEIRA,
R. F. M.; ABREU, M. L. T.; FERREIRA, A. S.;
SILVA, F. C. O.; HAESE, D. Exigência de lisina digestível para suínos machos castrados de alto potencial genético para deposição de carne magra na carcaça dos 15 aos 30 kg. Revista
Brasileira de Zootecnia, v. 35, n. 6, p. 2338-
2343, 2006.
ORLANDO, U.A.D. Nível de proteína bruta da ração e efeito da temperatura ambiente sobre o desempenho e parâmetros fisiológicos de leitoas em crescimento.Viçosa, MG:
Universidade Federal de Viçosa, 2001.
77p.Dissertação (Mestrado em Zootecnia) -
Universidade Federal de Viçosa, 2001.
PARR, T. M.; KERR, B. J.; BAKER, D. H. Isoleucine requirement of growing (25 to 45 kg) pigs.
Journal of animal science, v. 81, n. 3, p. 745-
752, 2003.
PARTRIDGE, I.G.; LOW, A.G.; KEAL, H.D. A note on the effect of feeding frequency on nitrogen use in growing boars given diets with varying levels of free lysine. Animal Production, v.40,n.2, p.375-
377, 1985.
PEDROZO, S. A. Níveis de lisina e relações treonina: lisina no desempenho e metabolismo de leitões desmamados. 2002.
142 f. Tese (Doutorado) - Universidade Federal do Rio Grande do Sul, Porto Alegre, 2002.
PENA, S. D. M.; LOPES, D. C.; ROSTAGNO, H. S.;
DE OLIVEIRA SILVA, F. C.; DONZELE, J. L.
Relações metionina mais cistina digestível: lisina digestível em dietas suplementadas com ractopamina para suínos em terminação. Revista
Brasileira de Zootecnia, v. 37, n. 11, p. 1978-
1983, 2008.
PEREIRA, A. A.; DONZELE, J. L.; DE OLIVEIRA, R.
F. M.; DE ABREU, M. L. T.; DE OLIVEIRA SILVA,
F. C.; DOS SANTOS MARTINS, M. Níveis de triptofano digestível em rações para suínos machos castrados de alto potencial genético na fase dos 97 aos 125 kg. Revista Brasileira de
Zootecnia, v.37, n.11, p.1984-1989, 2008.
POZZA, P. C.; GOMES, P. C.; DONZELE, J. L.;
FERREIRA, A. S.; LEÃO, M. I.; SANTOS, M. S.
D.; RODRIGUEIRO, R. J. B. Exigência de treonina para leitoas dos 15 aos 30 kg. Revista
Brasileira de Zootecnia, v. 29, n. 3, p. 817–822,
2000.
PRANDINI, A. L. D. O.; SIGOLO, S.;
MORLACCHINI, M.; GRILLI, E.; FIORENTINI, L.
Microencapsulated lysine and low-protein diets:
Effects on performance, carcass characteristics and nitrogen excretion in heavy growing–finishing pigs. Journal of animal science, v. 91, n. 9, p.
4226-4234, 2013.
PUPA, J.M.R.; ORLANDO, U.A.D.; DONZELE, J.L.
Requerimentos nutricionais de suínos nas condições brasileiras. In: WORKSHOP LATINO
AMERICANO BIOLATINA, NUTRIÇÃO DE AVES
E SUÍNOS, 1., 2001, Foz do Iguaçu. Anais...Foz do Iguaçu: 2001. p.143-153.
RODRIGUES, N. E. B.; DONZELE, J. L.; OLIVEIRA,
R. F. M. D.; LOPES, D. C.; FERREIRA, A. S.;
RODRIGUES FILHO, M.; ORLANDO, U. A. D.
Níveis de treonina em rações para leitoas com alto potencial genético para deposição de carne magra dos 30 aos 60 kg. Revista Brasileira de
Zootecnia, v. 30, n. 6, p. 2039-2045, 2001.
ROSSONI, M. C.; DONZELE, J. L.; SILVA, F. C. O.;
OLIVEIRA, R. F. M. D.; ABREU, M. L. L. T.;
SANTOS, F. D. A.; PEREIRA, C. M. C. Exigência de lisina digestível de fêmeas suínas selecionadas para deposição de carne magra, na carcaça dos 15 aos 30 kg. Revista Brasileira de
Saúde e Produção Animal, v.10, n.3, p.586-595,
2009.
SALYER, J. A.; TOKACH, M. D.; DEROUCHEY, J.
M.; DRITZ, S. S.; GOODBAND, R. D.; NELSSEN,
J. L. Effects of standardized ileal digestible tryptophan: lysine in diets containing 30% dried distillers grains with solubles on finishing pig performance and carcass traits. Journal of animal science, v. 91, n. 7, p. 3244-3252, 2013.
SARAIVA, E. P.; OLIVEIRA, R. F. M.; DONZELE, J. L.;
SILVA, F. C. O.; VAZ, R.; SIQUEIRA, J. C.;
OLIVEIRA, W. P. Níveis de treonina Digestivel em rações para leitoas dos 15 aos 30 kg mantidas em ambiente de alta temperatura. Revista Brasileira de Zootecnia, v. 35, n. 2, p. 485-490, 2006.
SERAO, M. C. R.; DONZELE, J. L.; SILVA, F.;
OLIVEIRA, R.; FERREIRA, A. S.; KILL, J. L.;
APOLÔNIO, L. R. Níveis de lisina digestível de fêmeas suínas selecionadas para deposição de carne magra na carcaça dos 30 aos 60 kg. Rev.
Bras. Saúde Prod. Anim., Salvador, v.13, n.2, p.433-443 abr./jun., 2012. ISSN 1519 9940.
SHEN, Y. B.; VOILQUE, G.; KIM, J. D.; ODLE, J.;
KIM, S. W. Effects of increasing tryptophan intake on growth and physiological changes in nursery pigs. Journal of animal science, v. 90, n. 7, p.
2264-2275, 2012.SOBRINHO, D. C. D. S.; DE OLIVEIRA JÚNIOR, G.
M.; RONER, M. N. B.; FERREIRA, A. S.; DE
OLIVEIRA, A. G.; DOS SANTOS, W. G.; DA
SILVA MORAIS, J. A. Lisina digestível para suínos machos castrados submetidos a estresse por calor dos 95 aos 115 kg. Revista Brasileira de Saúde e Produção Animal, v. 14, n. 3, 2013.
SOLBERG, J., BUTTERY, P.J., BOORMAN, K.N.
Effect of moderate methionine deficiency on food, protein and energy utilization in the chick. British
Poultry Science, v. 12,p.297-304, 1971.
TAVERNARI, F. DE. C. Atualização da proteína ideal para frangos de corte: valina e isoleucina. 2010. 75 f. Tese (Doutorado) -
Universidade Federal de Viçosa, Viçosa, 2010.
VIDAL, T. Z. B. Efeito da redução da proteína bruta e da suplementação de aminoácidos para suínos machos castrados, dos 70 aos 100 kg. Arq. Bras.
Med, v. 62, n. 4, p. 914-920, 2010.
VIEIRA VAZ, R. G. M.; OLIVEIRA, R.; DONZELE, J.
L.; FERREIRA, A. S.; SILVA, F.; KIEFER, C.;
RESENDE, W Exigência de aminoácidos sulfurados digestíveis para suínos machos castrados mantidos em ambiente de alta temperatura dos 15 aos 30 kg. Revista Brasileira
Zootecnia, v. 34, n. 5, p. 1633-1639, 2005.
VIEIRA VAZ, R. G. M.; OLIVEIRA, R.; DONZELE,
J. L.; FERREIRA, A. S.; SILVA, F.; KIEFER,
C.; RESENDE, W Exigência de aminoácidos sulfurados digestíveis para suínos machos castrados mantidos em ambiente de alta temperatura dos 15 aos 30 kg. Revista
Brasileira Zootecnia, v. 34, n. 5, p. 1633-
1639, 2005.
WAGUESPACK, A. M.; BIDNER, T. D.; PAYNE,
R. L.; SOUTHERN, L. L. Valine and isoleucine requirement of 20-to 45-kilogram pigs.
Journal of animal science, v. 90, n. 7, p.
2276-2284, 2012.
WALTON, M.J. Aspects of amino acid metabolism in teleost fish. In: COWEY, C.B.;
MACKIE, A.M.; BELL, J.G. Nutrition and feeding in fish. London: Academic Press,
1985, p. 47-67.
ZHU, C. L.; RADEMACHER, M.; DE LANGE, C.
F. M. Increasing dietary pectin level reduces utilization of digestible threonine intake, but not lysine intake, for body protein deposition in growing pigs. Journal of animal science, v.
83, n. 5, p. 1044-1053, 2005.










Soluções para Saúde Intestinal: produção sustentável de proteína animal e sucesso econômico
