Banco Ativo de Germoplasma de Capim-Elefante: Avaliação da Resistência à Cigarrinha-das-pastagens e Tolerância à Toxidez por Alumínio
Publicado: 20 de setembro de 2013
Por: Juarez Campolina Machado (Engenheiro Agrônomo, D.Sc. – Embrapa Gado de Leite); Carlos Eugênio Martins; Alexander Machado Auad; Wadson Sebastião Duarte da Rocha; Francisco José da Silva Lédo; Antonio Vander Pereira; Fausto de Souza Sobrinho; Flávio Rodrigo Gandolfi Benites.
Introdução
O capim-elefante (Pennisetum purpureum Schumach.) é considerado uma importante forrageira tropical devido à elevada produção de biomassa, fácil adaptação a diversos ecossistemas e boa aceitação pelos animais (LIRA et al., 2010). Pode ser utilizado sob as formas de capineira, pastejo, feno e silagem, e está entre as espécies forrageiras mais utilizadas para intensificação dos sistemas de produ ção de leite e carne (PEREIRA et al., 2010). Além do uso forrageiro, o capim-elefante tem sido proposto como uma alternativa sustentável para utilização da biomassa como insumo energético, sobretudo em função da sua alta eficiência fotossintética e elevada relação c arbono: nitrogênio (ANDERSON et al., 2008).
O Banco Ativo de Germoplasma de Capim-elefante da Embrapa (BAGCE), sob responsabilidade da Embrapa Gado de Leite, foi instituído no início da década de 80 e têm como finalidade a conservação, ampliação, valoração e uso da diversidade genética da espécie. A ampliação da variabilidade genética tem sido obtida por meio de coletas, introdução e intercâmbio com instituições nacionais e internacionais. Ao longo desses anos foram realizadas caracterizações e avaliações morfológica, agronômica e molecular dos acessos do BAGCE, de forma a permitir a compreensão da variabilidade existente na coleção. Contudo, é necessário ampliar a avaliação para estresses bióticos e abióticos, especialmente quanto à tolerância diferencial ao alumínio e a resistência à cigarrinha-das-pastagens.
Alguns trabalhos realizaram avaliações para esses estresses (MARTINS et al., 2010; AUAD et al., 2007), porém, normalmente, testaram um reduzido número de acessos, não representativos da variabilidade conservada. Pretende-se com essa publicação reunir as informações disponíveis, apresentar novas análises e direcionar os trabalhos futuros, haja vista que grande parte do germoplasma de capim-elefante conservado no BAGCE, foi avaliado conjuntamente.
Assim, o objetivo deste trabalho foi reunir as informações e complementar as avaliações e caracterizações já realizadas com avaliações da resistência à cigarrinha-das-pastagens e tolerância ao alumínio no germoplasma de capim-elefante e identificar acessos tolerantes e, ou resistentes a esses estresses.
Banco Ativo de Germoplasma de Capim-elefante
O Banco Ativo de Germoplasma de Capim-Elefante da Embrapa (BAGCE) integra a Plataforma Nacional de Recursos Genéticos, na Rede de Recursos Genéticos Vegetais (Rede Vegetal). O projeto da Rede Vegetal está estruturado em 14 projetos componentes sendo que o Projeto Componente 5 (Plantas Forrageiras) é coordenado pela Embrapa Gado de Corte, em colaboração com a Embrapa Recursos Genéticos e Biotecnologia. Em seu conjunto, os planos de ação abrangem os recursos genéticos de forrageiras específicas e os recursos genéticos autóctones regionais.
O BAGCE está sob a responsabilidade da Embrapa Gado de Leite, e desde a sua criação, vem conservando a variabilidade do gênero Pennisetum. Dentre as atividades desenvolvidas estão a conservação, ampliação, valoração e uso da diversidade genética.
O BAGCE possui 113 acessos, sendo: 94 de Pennisetum purpureum, nove raças cromossômicas (híbridos interespecíficos de P. purpureum x P. glaucum) e 10 acessos de espécies do pool gênico terciário do gênero (Pennisetum spp.). Dos 94 acessos de P. purpureum, 43 são originários de intercâmbio com outras instituições brasileiras, cinco da Colômbia, um da Costa Rica, quatro de Cuba, três da Índia, um do Panamá, um dos Estados Unidos e 36 são de origem desconhecida (Tabela 1). Também está sendo conservada uma coleção de trabalho de P. glaucum com 40 acessos.
A ampliação da variabilidade genética tem sido realizada por meio de coletas, introduções de outras instituições, intercâmbio com instituições internacionais e obtenção de novos híbridos interespecíficos. Até o momento foram realizadas caracterizações e avaliações morfológica (SHIMOYA et al., 2002), agronômica (SHIMOYA et al., 2001), e molecular (AZEVEDO et al., 2012) dos acessos.
As caracterizações e avaliações realizadas foram eficientes em discriminar os acessos e separá-los em grupos de relativa homogeneidade, de acordo com a caracterização realizada. Avaliações a respeito da fisiologia, resposta a estresses bióticos e abióticos e do potencial de utilização para produção de biomassa energética poderão ser realizadas para complementar as informações obtidas. Nesse sentido, a expectativa é promover o uso da variabilidade em Programas de Melhoramento e/ou Pré-melhoramento Genético de Capim-elefante.
Figura 1. Banco Ativo de Germoplasma de Capim-elefante da Embrapa. Créditos: Francisco José da Silva Lédo.

Tabela 1. Acessos do Banco Ativo de Germoplasma de Capimelefante da Embrapa.

Tolerância à toxidez por alumínio
A maioria dos solos brasileiros apresenta baixa fertilidade e problemas de acidez e toxidez por alumínio. A alta concentração de alumínio nos solos ácidos é, portanto, um dos principais fatores limitantes ao desenvolvimento das plantas (KOCHIAN et al., 2005) e assume papel importante na agricultura e pecuária nacionais, afetando diretamente os processos fisiológicos e metabólicos da grande maioria das espécies (HARTWIG et al., 2007).
O alumínio é um constituinte das partículas de argila do solo, e sua toxicidade é comum, na maioria dos solos onde o pH é suficientemente baixo (pH<5). Quando esse ponto é alcançado, parte do alumínio constituinte das partículas de argila migra para a fração trocável ou para a solução do solo. Esse elemento, nas camadas superficiais pode ser precipitado pela calagem, porém no subsolo permanece solúvel e tóxico às plantas (CAMAR GO & OLIVEIRA, 1981).
Existe ampla variabilidade no comportamento das plantas em condições de estresse causado pelo alumínio. No caso específico das f orrageiras, que normalmente ocupam áreas marginais, esses problemas são ainda mais graves (MARTINS et al., 2010). Entre as plantas forrageiras o capim-elefante é considerado como muito exigente, e de forma geral não se adapta a solos de baixa fertilidade e de elevada saturação por alumínio. Dentre as características negativas apresentadas pode-se citar o amarelecimento das folhas pela interferência na biossíntese da clorofila, arroxeamento das bainhas e margens do limbo, inibição do crescimento radicular e atrofiamento (S AMAC & TESFAYE, 2003).
A compreensão da variabilidade existente e a avaliação da tolerância ao alumínio tóxico no germoplasma de capim-elefante auxiliarão os programas de melhoramento e de r ecursos genéticos na identificação de acessos, que em combinações híbridas teriam maior chance de êxito na seleção para condições de estresse, potencializando o uso do germoplasma e possibilitando a obtenção de cultivares de capimelefante tolerantes à toxidez por alumínio.
Resistência à cigarrinha-daspastagens
A ação das cigarrinhas-das-pastagens é problema relevante na pecuária brasileira, sobretudo em função dos altos níveis populacionais, das ocorrências generalizadas e da severidade dos danos que causam.
O relacionamento inseto-planta engloba uma gama de espécies de cigarrinhas, associada a diversas espécies forrageiras, em diferentes sistemas de manejo e condições ecológicas (VALÉRIO, 2009). Entre as forrageiras, o capim-elefante é considerando como de baixa resistência, e em alguns casos, o ataque pode levar a planta à morte. A ação do inseto também torna a planta menos palatável, reduz o consumo pelos animais e consequentemente diminui a produção animal.
Danos econômicos foram registrados pelo ataque das espécies Notozulia entreriana, Deois schach, Mahanarva fimbriolata e Mahanarva spectabilis (TORRES E SILVA-TORRES, 2010). Entretanto, as cigarrinhas-das-pastagens do gênero Mahanarva são as de maior ocorrência e causadoras dos danos mais severos. O pico populacional das cigarrinhas coincide com a estação chuvosa do ano, quando as forrageiras estão em fase de maior desenvolvimento (VALÉRIO, 2009). Foi constatado também que elevadas densidades populacionais estão correlacionadas positivamente com umidade e temperatura. Contudo, para algumas espécies a ocorrência não está limitada ao período chuvoso (AUAD et al., 2009). Santos et al. (1995) registraram dois momentos de maior ataque do inseto, sendo o primeiro de outubro a novembro e o segundo em abril.
Apesar de existir um grande número de referências sobre a resistência à cigarrinha-das-pastagens em várias espécies forrageiras (THOMAS & LAPOINTE, 1989; VALÉRIO et al., 1997; CARDONA et al., 2004), às relativas ao capim-elefante são raras. Além disso, o controle químico é antieconômico; fazendo com que a praga se torne uma ameaça potencial.
Dessa forma, uma das principais demandas dos produtores em relação às instituições de pesquisa refere-se à obtenção de cultivares de capim-elefante resistentes às cigarrinhas-das-pastagens (AUAD et al., 2007). O uso de cultivar resistente é considerado o método ideal de controle, haja vista que as populações da praga podem ser reduzidas a níveis inferiores ao de dano econômico sem causar distúrbio ao ecossistema, além de ser de baixo custo e de fácil adoção.
Estudos têm constatado que a resistência à cigarrinha está sob controle genético, e pode ser facilmente manipulada em um programa de melhoramento de plantas (MILES et al., 1995; SOUZA SOBRINHO, 2005). Considerando a diversidade genética existente no germoplasma de capim-elefante, acredita ser possível identificar acessos com resistência às cigarrinhas, que poderão ser usadas como fonte de alelos favoráveis nos programas de melhoramento ou de recursos genéticos de capim-elefante, potencializando o uso do germoplasma.
Material e Métodos
Tolerância à toxidez por alumínio
Os experimentos foram instalados em casa de vegetação no delineamento inteiramente casualizado, com três repetições nos anos de 2007 e 2011, e com quatro repetições no ano de 2009, sendo que foram avaliados 15 acessos em 2007 e 2009 e 44 acessos em 2011.
As parcelas experimentais foram constituídas de um vaso com uma planta, as quais foram propagadas por estacas em bandejas de plástico com substrato comercial. Após o per íodo de enraizamento, as plântulas foram selecionadas quanto à uniformidade de tamanho de parte aérea e de raízes, e transplantadas para vasos plásticos, com volume de dois litros de solução nutritiva de Clark (1975).
Durante os primeiros sete dias as plantas cresceram em solução meia força, ou seja, com metade da concentração de nutrientes recomendada, sem aplicação de alumínio. Depois desta etapa e durante 29 dias, promoveu-se, semanalmente, a troca da solução nutritiva, acrescida de 15 mg/L de alumínio, nível considerado o mais adequado para identificar materiais tolerantes e sensíveis à toxidez por alumínio (MARTINS et al. 2010). Em intervalos de três dias procedia-se também o rodízio no posicionamento dos vasos nas bancadas para assegurar a homogeneidade das condições ambientais.
As características avaliadas foram: peso seco de parte aérea (PSPA) e peso seco de raiz (PSR), medidas em gramas (g). Inicialmente foram realizadas análises de variância (teste F) para as características avaliadas, considerando o efeito de tratamentos como fixo. Constatada diferenças significativas, foi realizado o teste de média de Scott & Knott a 5% de probabilidade.
Figura 2. Avaliações da tolerância à toxidez por alumínio. Créditos: Wadson Sebastião Duarte da Rocha, Carlos Eugênio Martins e Juarez Campolina Machado.

Resistência à cigarrinha-das-pastagens
Os experimentos foram conduzidos em casa de vegetação (temperatura mínima, média e máxima de 20 ºC, 29 ºC e 36 ºC, respectivamente), nos anos de 2007, 2009 e 2012 a partir de criação de M. spectabilis da Embrapa Gado de Leite, que serviu de suprimento de material vivo para os ensaios.
Adultos do inseto foram direcionados ao Laboratório de Entomologia, sexados e mantidos em gaiolas cilíndricas de acrílico para a produção de ovos. Em cada gaiola foi alocada uma planta de capim-elefante com a base envolvida por gaze umedecida em água destilada, que serviu de substrato para oviposição. Para a retirada dos ovos, a gaze foi colocada sobre um conjunto de peneiras e submetido à água corrente, de forma que os ovos ficavam retidos na peneira mais fi na (400 mesh de abertura).
Posteriormente, os ovos obtidos foram colocados em placas de Petri de 5 cm de diâmetro, forradas com papel-filtro e mantidas em câma - ra climatizadas (28 ± 2 ºC, 14 horas de fotofase e umidade relativa de 70 ± 10%) até próximo à eclosão.
O estudo se baseou no mecanismo da resistência por antibiose. Estabeleceram- se plantas de cada acesso, sendo uma por repetição, a partir de estacas de um nó, em vasos de 500 mL, contendo uma mistura de substrato de solo, areia e esterco (1:1:1). Após o estabelecimento, as plantas foram infestadas. Seis ovos, previamente incubados em laboratório e próximos à eclosão foram depositados em cada um dos acessos de capim-elefante, incluindo as testemunhas, Roxo de Botucatu (suscetível) e Pioneiro (resistente). As raízes das plantas foram expostas para facilitar a alimentação das ninfas do inseto-praga.
Quarenta dias após a deposição dos ovos, foi realizada a contagem das ninfas de terceiro, quarto e quinto ínstares, bem como o número total de ninfas sobreviventes, nos diferentes acessos de capim-elefante.
O delineamento experimental foi inteiramente casualizado, com dez repetições no ano de 2007 (25 acessos avaliados), oito repetições no ano de 2009 (24 acessos avaliados) e cinco repetições no ano de 2012 (90 acessos avaliados). Os dados foram transformados em (x + 1) 0,5 e posteriormente foi realizada a análise de variância (teste F), considerando o efeito de tratamentos como fixo. Constatada diferenças significativas, foi realizado o teste de média de Scott & Knott a 5% de probabilidade.
Para todas as análises estatísticas utilizaram-se os recursos computacionais do Programa GENES (CRUZ, 2006).
Figura 3. Avaliação da resistência à cigarrinha-das-pastagens. Créditos: Alexander Machado Auad.

Resultados e Discussão
Tolerância à toxidez por alumínio
Os resultados do desempenho médio dos acessos de capim-elefante em relação ao peso seco da raiz (PSR) e da parte aérea (PSPA) estão apresentados nas Figuras 4, 5 e 6. A identificação dos acessos promissores foi realizada considerando aqueles que se destacaram individualmente em cada um dos experimentos (ano de avaliação).
Foram constatadas diferenças significativas (P<0,05) entre os acessos para características mensuradas, nos diferentes ensaios, exceto para peso seco da parte aérea, no experimento realizado no ano de 2007, indicando a existência de variabilidade genética no germoplasma de capim-elefante para a tolerância à toxidez por alumínio em solução nutritiva. Resultados semelhantes também foram obtidos na avaliação de outras forrageiras tropicais (ALMEIDA et al., 2000; BITENCOURT et al., 2011; MIGUEL et al., 2011), sugerindo viabilidade no melhoramento genético e possibilidade de identificação de genes/alelos de interesse para essa característica.
Figura 4. Médias do peso seco da parte aérea (PSPA) e do peso seco das raízes (PSR) em gramas (g), de acessos de capim-elefante avaliados em solução nutritiva. Médias seguidas de mesma letra maiúscula para o PSPA e minúscula para o PSR, pertencem ao mesmo grupo pelo teste de Scott-Knott ao nível de5%de probabilidade. Juiz de Fora-MG, 2007.
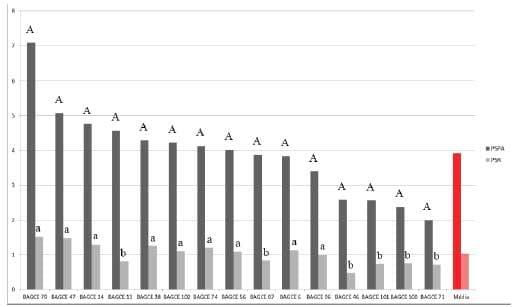
Os coeficientes de variação (CV) foram considerados de baixos a médios, exceto para a característica PSPA, no experimento realizado no ano de 2007 (CV=30%). Provavelmente por esse motivo não foram detectadas diferenças significativas nesse caso.
Para o PSPA, no experimento realizado em 2009, os acessos foram separados em três grupos distintos, com destaque para o BAGCE 92 que obteve a maior média. No ano de 2011, os acessos foram separados também em três grupos, sendo que os acessos BAGCE 53, BAGCE 68, BAGCE 1, BAGCE 2, BAGCE 54, BAGCE 7, BAGCE 55, BAGCE 33, BAGCE 17, BAGCE 37, BAGCE 51, BAGCE 62, BAGCE 11, BAGCE 61, BAGCE 26, BAGCE 18, BAGCE 45, BAGCE 28, BAGCE 29, BAGCE 35 e BAGCE 59 formaram o grupo superior.
Figura 5. Médias do peso seco da parte aérea (PSPA) e do peso seco das raízes (PSR) em gramas (g), de acessos de capim-elefante avaliados em solução nutritiva. Médias seguidas de mesma letra maiúscula para o PSPA e minúscula para o PSR, pertencem ao mesmo grupo pelo teste de Scott-Knott ao nível de5%de probabilidade. Juiz de Fora-MG, 2009.
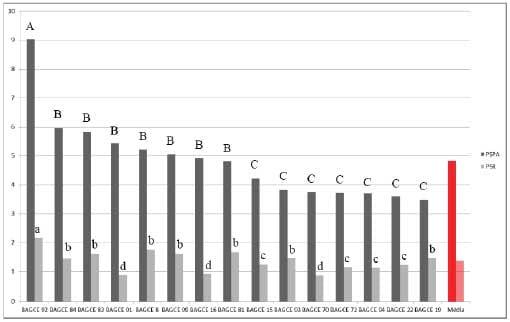
Para o PSR, no ano de 2007, constatou-se a separação dos acessos em dois grupos, com o grupo de melhor desempenho formado por BAGCE 79, BAGCE 47, BAGCE 14, BAGCE 38, BAGCE 74, BAGCE 6, BAGCE 102, BAGCE 56, BAGCE 96. Em 2009, constatou-se a formação de quatro grupos distintos, com o acesso BAGCE 92 se destacando dos demais. Para o experimento realizado em 2011, novamente os acessos foram separados em quatro grupos sendo que BAGCE 53, BAGCE 68, BAGCE 7 e BAGCE 55 formaram o grupo de melhor desempenho.
Considerando as duas características conjuntamente os acessos BAGCE 79 e BAGCE 47 for am os únicos classificados no grupo com desempenho superior para ambas as características (Figura 4), no ano de 2007. No ano de 2009 apenas o acesso BAGCE 92 se destacou (Figura 5). Em 2011 os mais promissores foram BAGCE 53, BAGCE 68, BAGCE 7 e BAGCE 55 (Figura 6). Esses acessos poderão ser utilizados em programas de pré-melhoramento e melhoramento genético como fonte de alelos visando tolerância à toxidez por alumínio.
Figura 6. Médias do peso seco da parte aérea (PSPA) e do peso seco das raízes (PSR) em gramas (g), de acessos de capim-elefante avaliados em solução nutritiva. Médias seguidas de mesma letra maiúscula para o PSPA e minúscula para o PSR, pertencem ao mesmo grupo pelo teste de Scott-Knott ao nível de5%de probabilidade. Juiz de Fora-MG, 2011.
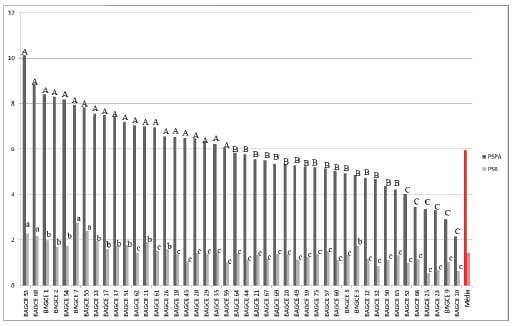
Os resultados obtidos nesse trabalho evidenciam ampla variação para as características, confirmando a existência de variabilidade genética entre os acessos de capim-elefante. Além disso, permitem antever a possibilidade de sucesso com a seleção de genótipos d e capim-elefante tolerantes ao alumínio.
Vale ressaltar que os experimentos foram conduzidos em casa de vegetação, utilizando solução nutritiva. Isto é um indicativo da tolerância a solos ácidos, contudo faz-se necessário a confirmação dess es resultados com a realização de experimentos de campo (MARTINS et al., 2010). Nesse caso, o número de acessos testados pode ser menor, sendo a sua escolha baseada nos resultados desse trabalho.
Resistência à cigarrinha-das-pastagens
Foram constatadas diferenças significativas (P<0,05) par a a sobrevivência ninfal de M. spectabilis em todos os experimentos, sugerindo a existência de variabilidade genética para a resistência e suscetibilidade a esse inseto-praga no germoplasma de capim-elefante, por meio do mecanismo de antibiose.
A sobrevivência ninfal nos acessos testados variou de 33% a 100 %, e as médias foram separadas em grupos distintos (Figuras 7, 8 e 9), contudo os níveis de sobrevivência obtidos são ainda considerados altos. De acordo com Cardona et al. (1999), para espécies de Brachiaria, somente genótipos com sobrevivência ninfal inferior a 30% deveriam ser considerados resistentes ao inseto-praga.
Figura 7. Sobrevivência média de ninfas de terceiro, quarto e quinto ínstares (%) de Mahanarva spectabilis em acessos do Banco Ativo de Germoplasma de Capim-elefante da Embrapa. Médias seguidas de mesma letra pertencem ao mesmo grupo pelo teste de Scott-Knott ao nível de 5% de probabilidade. Juiz de Fora-MG, 2007.
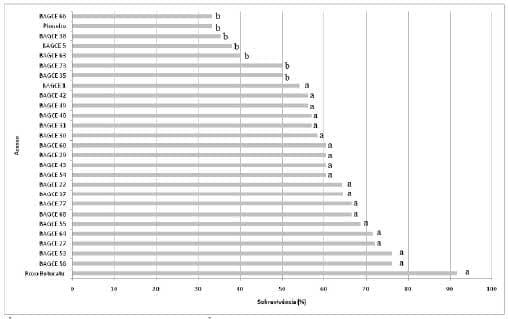
O acesso Roxo Botucatu esteve sempre no grupo de maior sobrevivência ninfal, já a cultivar Pioneiro se destacou pela menor sobrevivência de ninfas nos diferentes experimentos. Esses genótipos s ão, respectivamente, candidatos a testemunhas suscetível e resistente, pelo mecanismo de antibiose.
Figura 8. Sobrevivência média de ninfas de terceiro, quarto e quinto ínstares (%) de Mahanarva spectabili em acessos do Banco Ativo de Germoplasma de Capim-elefante da Embrapa. Médias seguidas de mesma letra pertencem ao mesmo grupo pelo teste de Scott-Knott ao nível de 5% de probabilidade. Juiz de Fora-MG, 2009.
Foram agrupados conjuntamente à cultivar resistente, seis acessos no ano de 2007, vinte acessos no ano de 2009 e vinte e nove acesso no ano de 2012 (Figuras 7, 8 e 9). Nos demais acessos avaliados a porcentagem de sobrevivência de M. spectabilis não diferiu estatisticamente da testemunha suscetível Roxo Botucatu, com sobrevivência média de ninfas no grupo de 65%, 78% e 70%, nos anos de 2007, 2009 e 2012 respectivamente, caracterizando reação de suscetibilidade.
No presente estudo, grande parte (95%) dos acessos conservados no Banco Ativo de Germoplasma de Capim-elefante da Embrapa (BAGCE) foram avaliados. Considerando que o BAGCE é uma coleção representativa da variabilidade genética da espécie, ressalta-se que mesmo os acessos considerados promissores, ainda apresentam alta sobrevivência ninfal (>30%) (Figuras 7, 8 e 9), o que pode limitar a expansão dessa espécie forrageira em áreas onde o inseto-praga tem ocasionado danos econômicos. Dessa forma, novas alternativas devem ser utilizadas para o desenvolvimento de cultivares resistentes.
Vale ressaltar que os resultados apresentados referem-se a contagem das ninfas de terceiro, quarto e quinto ínstares. Esse critério para identificação de resistência por antibiose é apoiado pelo fato de ninfas com 45 dias de idade ainda não atingiram o terceiro instar, indicando que a planta está promovendo condição desfavorável ao desenvolvimento do inseto. Contudo quando se considerou o número total de ninfas sobreviventes, os resultados se mantiveram concordantes, haja vista que a correlação entre essas duas variáveis nos diferentes acessos de capim-elefante foi de r=0,88 (P<0,01).
Os resultados deste trabalho corroboram o relatado por Souza Sobrinho (2005), o qual comenta que a suscetibilidade à cigarrinha-das- -pastagens é o principal fator limitante a ampla utilização de determinadas espécies forrageiras e que estratégias de melhoramento visando a resistência, e o manejo do inseto-praga deverão ser priorizados pela pesquisa.
Dentre os acessos testados, BAGCE 63, BAGCE 5, BAGCE 69, BAGCE 51, BAGCE 6, BAGCE 52 e BAGCE 13 se destacaram em pelo menos dois ensaios (Figuras 7, 8 e 9), com moderada sobrevivência de ninfas de M. spectabilis. Esses acessos, juntamente com a testemunha resistente (Pioneiro) poderão ser utilizados no programa de melhoramento do capim-elefante como estratégia para a obtenção de cultivares resistentes ao inseto, contudo outras possibilidades deverão também ser utilizadas.
Figura 9. Sobrevivência média de ninfas de terceiro, quarto e quinto ínstares (%) de Mahanarva spectabilisem acessos do Banco Ativo de Germoplasma de Capim-elefante da Embrapa.

Uma alternativa seria avaliar espécies relacionadas (Pennisetum spp.) visando identificar genes/alelos de resistência e posterior incorporação em programas de pré-melhoramento. Outra opção seria avaliar os compostos liberados pelas plantas em resposta ao ataque dos insetos que influenciariam na resistência (HEIL, 2008), e utilizar como instrumento de seleção.
É importante comentar que os resultados obtidos neste experimento envolvendo M. spectabilis devem ser confirmados por testes de campo, possivelmente abrangendo outras espécies de cigarrinhas-das-pastagens.
Conclusões
Existe variabilidade genética entre os acessos de capim-elefante para tolerância à toxidez por alumínio e para a resistência e suscetibilidade à cigarrinha-das-pastagens.
Para a toxidez por alumínio o nível de tolerância identificado nos acessos foi considerado satisfatório.
Para a resistência a cigarrinha-das-pastagens, os níveis de sobrevivência ninfal obtidos foram considerados altos, e novas estratégias devem ser utilizadas para o desenvolvimento de cultivares resistentes ao inseto-praga.
Os acessos BAGCE 79, BAGCE 47, BAGCE 92, BAGCE 53, BAGCE 68, BAGCE 7 e BAGCE 55 são promissores como fonte de alelos visando o melhoramento para condições de estresse por alumínio.
Referências
ALMEIDA, A. A. S.; MONTEIRO, F. A.; JANK, L. Avaliação de Panicum maximum JACQ. para tolerância ao alumínio em solução nutritiva. Revista Brasileira de Ciência do Solo, v. 24, p. 339-344, 2000.
ANDERSON, W.; CASLER, M.; BALDWIN, B. Improvement of perennial forage species as feedstock for bioenergy. In: VERMERRIS, W. (Ed.). Genetic improvement of bioenergy crops. Springer, 2008. p. 308-345.
AUAD, A. M.; CARVALHO, C. A.; SILVA, D. M.; DERESZ, F. Flutuação populacional de cigarrinhas-das-pastagens em braquiária e capim-elefante. Pesquisa Agropecuária Brasileira, v. 44, n. 9, p. 1205-1208, 2009.
AUAD, A. M.; SIMÕES, A. D.; PEREIRA, A. V.; BRAGA, A. L. F.; SOUZA SOBRINHO, F.; LÉDO, F. J. da S ., PAULA-MORAES, S. V.; OLIVEIRA, S. A.; FERREIRA, R. B. Seleção de genótipos de capim- -elefante quanto à resistência à cigarrinha-das-pastagens. Pesquisa Agropecuária Brasileira, v. 42, p. 1077-1081, 2007.
AZEVEDO, A. L. S.; COSTA, P. P. ; MACHADO, J. C. ; MACHADO, M. A.; PEREIRA, A. V.; LÉDO, F. J. da S . Cross species amplification of Pennisetum glaucum microsatellite markers in Pennisetum purpureum and genetic diversity of napier grass accessions. Crop Science, v. 52, p. 1776-1785, 2012.
BITENCOURT, G. A.; CHIARI, L.; LAURA, V. A.; VALLE, C. B.; JANK, L.; MORO, J. R. Aluminum tolerance on genotypes of signal grass.. Revista Brasileira de Zootecnia. v. 40, n. 2, 2011.
CARDONA, C.; FORY, P.; SOTELO, G.; PABON, A.; DIAZ, G.; MILES, J. W. Antibiosis and tolerance to five species of spittlebug (Homoptera: Cercopidae) in Brachiaria spp.: implications for breeding for resistance. Journal of Economic Entomology, v. 97, p. 635-645, 2004.
CARDONA, C.; MILES, J. W.; SOTELO, G. An improved methodology for massive screening of Brachiaria spp. genotypes for resistance to Aeneolamia varia (Homoptera: Cercopidae). Journal of Economic Entomology. v. 92, n. 2, p. 490-496, 1999.
CAMARGO, C. E. O.; OLIVEIRA, O. F. Tolerância de cultivares de trigo a diferentes níveis de alumínio em solução nutritiva e no solo. Bragantia, Campinas, v. 40, p. 21-23, 1981.
CLARK, R. B. Characterization of phosphatase in intact maize roots. Journal of Agricultural and Food Chemistry, v. 23, p. 458-460, 1975.
CRUZ, C. D. Programa GENES: estatística experimental e matrizes. Viçosa: UFV, 2006. 285 p.
HARTWIG, I.; OLIVEIRA, A. C.; CARVALHO, F. I. F.; BERTAN, I.; SILVA, J. A. G.; SCHMIDT, D. A. M.; VALERIO, I. P.; MAIA, L. C.; FONSECA, D. A. R.; REIS, C. E. S. Mecanismos associados à tolerância ao alumínio em plantas. Semina: Ciências Agrárias; v. 28, p. 219-228, 2007.
HEIL, M. Indirect defense via tritrophic interactions. New Phytologist, v. 178, p. 41-61, 2008.
KOCHIAN, L. V. PINEROS, M. A. HOEKENGA, O. A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant and Soil, v. 274, n. 1, p. 175-195, 2005.
LIRA, M. A.; SANTOS, M. V. F.; DUBEUX JÚNIOR, J. C. B; MELLO, A. C. L (Eds.). Capim-elefante: fundamentos e perspectivas. Recife: IPA/ UFRPE, 2010. 229 p.
MARTINS, C. E.; SOUZA SOBRINHO, F.; GOMES, F. T.; ROCHA, W. S. D.; BRIGHENTI, A. M. Tolerância à toxidez por alumínio em capim-elefante. Pesquisa Aplicada & Agrotecnologia. v. 3, n. 1, p. 95-101, 2010.
MIGUEL, P. S. B.; ROCHA, W. S. D.; SOUZA SOBRINHO, F.; MARTINS, C. E.; GOMES, F. T.; OLIVEIRA, A. V. de; CARVALHO, C. A. de. Seleção de genótipos de Brachiaria ruziziensis quanto ao alumínio em solução nutritiva. II: Avaliação da tolerância ao alumínio. Revista de Ciências Agrárias, v. 34, n. 1, p. 163-172, 2011.
MILES, J. W.; LAPOINTE, S. L.; ESCANDON, M. L.; SOTELO, G. Inheritance of resistance to spittle bug (Homoptera: Cercopidae) in Inter specific Brachiaria spp. hybrids. Journal of Economic Entomology, v. 88, n. 5, p. 1477-1481, 1995.
PEREIRA, A. V.; AUAD, A. M., LÉDO, F. J. S.; BARBOSA, S. Pennisetum purpureum. In: FONSECA, D. M., MARTUSCELLO, J. A. (Eds.). Plantas Forrageiras. Viçosa: UFV, 2010. p. 198-220.
SAMAC, D. A.; TESFAYE, M. Plant improvement for tolerance to aluminium in acid soils - a review. Plant Cell, Tissue and Organ Culture, v. 75, p. 189–207, 2003.
SANTOS, A. V. dos; VALERIO, J. R.; SOUZA, A. P. de; OLIVEIRA, M. C. M. Níveis populacionais de ninfas de Mahanarva frimbriolata (Stal, 1854) (Homoptera: Cercopidae) em cultivares de capim-elefante, Pennisetum purpureum. In: CONGRESSO BRASILEIRO DE ENTOMOLOGIA, 15., 1995, Caxambú. Resumos... Belo Horizonte: SEB/ESAL, 1995. p. 761.
SHIMOYA, A.; CRUZ, C. D.; FERREIRA, R. P.; PEREIRA, A. V.; CARNEIRO, P. C. S. Divergência genética entre acessos de um banco de germoplasma de capim-elefante. Pesquisa Agropecuária Brasileira, Brasília, v. 37, n. 7, p. 971-980, 2002.
SHIMOYA, A.; FERREIRA, R. de P.; PEREIRA, A. V.; CRUZ, C. D.; CARNEIRO, P. C. S. Comportamento morfo-agronômico de genótipos de capim-elefante. Revista Ceres, Viçosa, v. 48, n. 276, p. 1-19, 2001.
SOUZA SOBRINHO, F. Melhoramento de forrageiras no Brasil. In: EVANGELISTA, A. R.; AMARAL, P. N. C.; PADOVANI, R. F.; TAVARES, V. B.; SALVADOR, F. M.; PERÓN, A. J. (Eds.). Forragicultura e pastagens: temas em evidência. Lavras: UFLA, 2005. p. 65-120.
TORRES, J. B.; SILVA-TORRES, C. S. A. Manejo de insetos pragas do capim- elefante (Pennisetum purpureum Schum.). In: LIRA, M. A.; SANTOS, M. V. F.; DUBEUX JÚNIOR, J. C. B; MELLO, A. C. L (Eds.). Capim-elefante: fundamentos e perspectivas. Recife: IPA/UFRPE, 2010. p. 87-110.
THOMAS, D.; LAPOINTE, S. Testing new accessions of guinea grass (Panicum maximum Jacq.) for acid-soils and resistance to spittlebug (Aeneolamia reducta).Tropical Grasslands, St. Lucia, v. 23, n. 4, p. 232-239, 1989.
VALÉRIO, J. R.; JELLER, H.; PEIXER, J. Seleção de introduções do gênero Brachiaria (Griseb) resistentes à cigarrinha Zulia entreriana (Berg) (Homoptera: Cercopidae). In: Anais da Sociedade Entomológica do Brasil, v. 26, p. 383-387, 1997.
VALÉRIO, J. R. Cigarrinha-das-pastagens. Campo Grande: Embrapa Gado de Corte, 2009. 51 p. (Embrapa Gado de Corte. Documentos, 179).
***O trabalho foi originalmente publicado por Empresa Brasileira de Pesquisa Agropecuária -Embrapa Gado de Leite / Ministério da Agricultura, Pecuária e Abastecimento - ISSN 1516-7453 Dezembro/2012.
Autores:

Embrapa
Embrapa
Embrapa
Embrapa
Embrapa
Mostrar mais
Recomendar
Comentário
Compartilhar

Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.

