2 Pós-Doc ESALQ-USP, bolsista FAPESP. 3 Departamento de Zootecnia - UFV. 4 ESALQ-USP. 5 UNIP - Jacareí - SP. 6 Embrapa Gado de Leite, Juiz de Fora – MG. 7 Zootecnista. * Pesquisador Bolsista CNPq.
Introdução
Não é recente a intenção de alterar a composição do leite, principalmente em relação à gordura. A perspectiva de manipular a gordura do leite visa atender à demanda de um mercado consumidor cada vez mais exigente em relação ao consumo de determinadas gorduras saturadas, em razão de seus efeitos deletérios sobre a saúde humana.
Uma das características do leite bovino é a grande proporção de ácidos graxos (AG) saturados, com cadeias de 4 a 16 carbonos, resultantes da síntese de novo. Alguns destes AG são apontados como precursores do colesterol sangüíneo de baixa densidade (LDL), responsável por doenças cardiovasculares (Parodi, 1999). Entretanto, é interessante aumentar a participação de ácidos graxos de cadeia longa, mono e poliinsaturados, na composição da gordura do leite, pois estes AG possibilitam redução da incidência de doenças coronarianas, com o aumento do colesterol de alta densidade, o HDL (Demeyer & Doreau, 1999).
Extensas revisões são encontradas na literatura relatando a dificuldade da manipulação e o incremento de determinados AG no leite (Chilliard et al., 2000). Segundo Demeyer & Doreau (1999), a gordura do leite é originada a partir dos lipídios sintetizados na glândula mamária (40 a 50%) e dos ácidos graxos pré-formados absorvidos da corrente sangüínea. Aproximadamente 10% dos AG circulantes têm origem na mobilização dos lipídios corpóreos, enquanto o restante é de origem dietética. No início da lactação, a extensão do balanço energético negativo pode determinar maior secreção de AG de cadeia longa no leite (Grummer, 1991).
Existe alta correlação entre os AG do leite e os da digesta duodenal, entretanto, a correlação entre os AG dietéticos e os do leite é baixa. São três os fatores que mais modificam os AG da dieta em relação aos absorvidos no duodeno (Chilliard et al., 2000): primeiro, a população microbiana age sobre os AG insaturados, promovendo a saturação, em uma rota denominada bio-hidrogenação ruminal; segundo, a microbiota ruminal sintetiza ácidos graxos de forma similar à síntese de novo; finalmente, a Δ9- desaturase age nos enterócitos e na glândula mamária, que inclui uma ligação dupla cis-9 nos AG, por exemplo, transformando o ácido esteárico (C18:0) em oléico (cis-9 C18:1) e o vacênico (trans-11 C18:1) em CLA cis-9 trans-11 C18:2.
A fase da lactação, o balanço energético, o nível de alimentação, a proporção de concentrado, o pH do rúmen e a utilização de tampões e ionóforos são fatores que influenciam o perfil da gordura do leite (Palmquist et al., 1993). No entanto, a inclusão de lipídios na dieta provoca as maiores alterações na gordura do leite, que são dependentes do nível e do tipo de lipídio e da extensão da bio-hidrogenação ruminal (Grummer, 1991).
Em experimentos in vitro, os ionóforos têm propiciado aumentos na concentração de isômeros trans-C18:1 e redução nas taxas de hidrólise dos ácidos graxos, assim como na velocidade e na extensão da bio-hidrogenação (Fellner et al., 1997). Jenkins et al. (2003) observaram, em fermentadores contínuos, que, além de aumentar todos os isômeros trans-C18:1, a combinação monensina e óleo de soja produziu efeitos aditivos em relação a trans-10 C18:1.
Griinari et al. (1998) demonstraram que a bio-hidrogenação incompleta esteve associada a alterações no ambiente ruminal, provocadas por dietas com baixo teor de fibra e alto de concentrado, que incidem em aumento proporcional na concentração de trans-10 C18:1, relacionada à redução no teor de gordura do leite.
São raros os trabalhos que envolvem efeitos de ionóforos e gorduras sobre o perfil de ácidos graxos do leite. Cant et al. (1997) utilizaram monensina e óleo de peixe e Johnson et al. (1988) associaram lasalocida com amendoim em grão como fonte de gordura. Em ambos os trabalhos, a proporção dos AG insaturados foi aumentada e o teor de gordura foi reduzido, enquanto o perfil dos isômeros trans-C18:1 não foi avaliado.
Os autores supracitados sugeriram que a associação de monensina e óleos vegetais reduz o teor de gordura do leite, em decorrência das alterações da população microbiana provocadas pelo ionóforo, além de possibilitar aumentos na concentração CLA (Ácido Linoléico Conjugado). De modo, objetivou-se avaliar o perfil de ácidos graxos, incluindo os isômeros trans-C18:1, e o conteúdo de CLA no leite de vacas alimentadas por longo período com a combinação monensina e óleo de soja na dieta.
Material e Métodos
O experimento foi conduzido no Campo Experimental de Coronel Pacheco – EMBRAPA Gado de Leite, no período de maio a agosto de 2002. As análises laboratoriais foram realizadas no Laboratório de Nutrição Animal do Departamento de Zootecnia da Universidade Federal de Viçosa e no Laboratório de Nutrição e Crescimento Animal da ESALQ/USP.
Foram utilizadas 16 vacas 7/8 Holandês-Gir, multíparas, com peso inicial de 472±41 kg e 29,9±9 dias em lactação. O delineamento experimental foi em blocos ao acaso, em um arranjo fatorial 2 x 2: que constitui na presença ou não de monensina sódica (33 ppm) e de óleo de soja (3,9%) na dieta. Os tratamentos foram conmpostos das dietas: CT (controle) = sem monensina ou óleo; MN = com monensina; OL = com óleo de soja; e OM = com óleo e monensina.
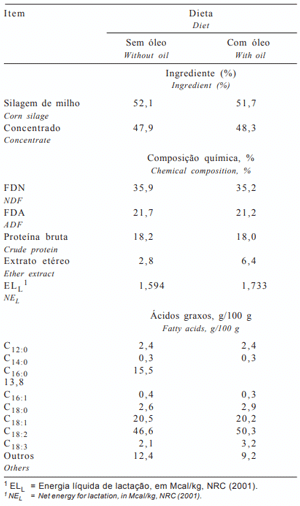
Os animais foram alimentados com 52% de silagem de milho e 48% de concentrado, conforme descrito na Tabela 1. As dietas foram formuladas para atender ou exceder as recomendações do NRC (2001) para vacas com produção de 25 kg/dia de leite com 3,5% de gordura. Os animais foram mantidos confinados, durante 84 dias, em free-stall, com cochos individuais regulados eletronicamente em um sistema tipo “Calan Gate” (Calan Data Ranger®), sendo alimentados e ordenhados duas vezes ao dia.
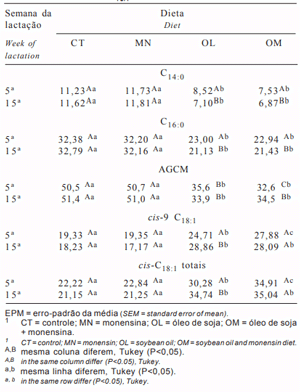
Tabela 1 - Ingredientes, composição química e perfil dos ácidos graxos das dietas experimentais
As amostras de leite para a avaliação do perfil dos ácidos graxos foram coletadas individualmente em duas ordenhas consecutivas na 5a e 15a semanas da lactação. Foram retiradas alíquotas compostas de 2% da ordenhas da manhã e da tarde; uma porção foi encaminhada para a análise por espectrofotometria (Laboratório de Qualidade do Leite, Embrapa Gado de Leite) e o restante, para a extração e metilação da gordura.
A extração da gordura foi realizada pelo método descrito por Nourooz-Zadeh & Appelqvist (1988). Foram transferidos 17 mL de leite (em duplicata) para um funil de separação, com acréscimo de 30 mL de isopropanol. Após agitação, foram acrescentados 22,5 mL de hexano, agitando-se o líquido por mais três minutos. A mistura foi centrifugada a 2.520 g, por 5 minutos, a 5ºC. A camada superior foi transferida para outro funil de separação e a inferior, extraída mais duas vezes com 22,5 mL de hexano e esses extratos foram adicionados ao primeiro. A água foi retirada dos extratos com a adição de 15 mL de Na2SO4 0,47 M. A camada de hexano foi coletada em frasco e evaporada a 50ºC em evaporador rotativo com fluxo de nitrogênio contínuo. O resíduo foi dissolvido em 50 mL de clorofórmio e seco com Na2SO4 anidro. Aproximadamente 40 mg do resíduo foram utilizados para a formação dos ésteres metílicos, com o uso de uma solução de metóxido de sódio em metanol, conforme descrito por Christie (1982), e analisados por cromatografia gasosa.
Os ésteres metilados dos ácidos graxos foram separados em uma coluna capilar de 100 m de sílica fundida (SP-2560) com hidrogênio como gás de arraste (1,8 mL/min) e detector de ionização de chama (FID). Cada amostra foi rodada como descrito por Griinari et al. (1998), com um gradiente de temperatura de 70 a 240°C para determinação dos picos de identificação dos AG.
Após a identificação dos picos, uma manteiga padrão (CRM 164; Commission of the European Communities, Community Bureau of Reference, Brussels, Belgium) foi utilizada para a certificação da recuperação dos ácidos graxos de acordo com os picos e tempos de retenção. A concentração dos ácidos graxos foi expressa em g/100 g dos ácidos graxos totais recuperados.
O delineamento experimental foi em blocos ao acaso com um período de avaliação de 12 semanas, em que o perfil dos ácidos graxos foi comparado em duas fases da lactação: na 5a e 15a semanas pós-parto. A análise estatística foi realizada utilizando-se o comando Proc GLM do programa SAS (1999). Para a comparação dos ácidos graxos em relação aos tratamentos, foram incluídos no modelo os efeitos das dietas, dos blocos e da fase. Utilizou-se a interação oleo*mon*bloco como erro ‘B’ para comparação dos ácidos graxos em relação à semana de lactação. Nas tabelas, os efeitos principais são reportados com os níveis de significância menores que 5%, exceto quando mencionados por algum interesse na discussão.
Resultados e Discussão
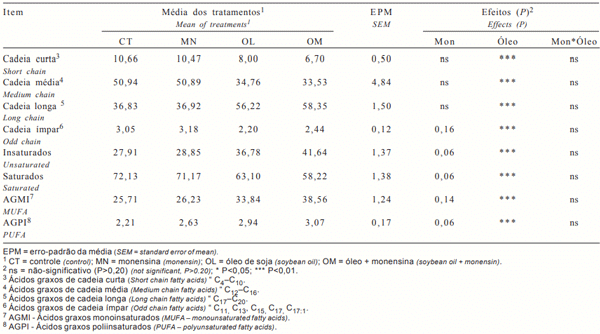
A análise cromatográfica apresentou recuperação de 97,8% dos ácidos graxos do leite amostrado, descritos nas tabelas a seguir em g/100 g do total de ácidos graxos recuperados. Na Tabela 2 são apresentados os grupos dos principais ácidos graxos do leite e os efeitos dos tratamentos e, na Tabela 3, são detalhados estes ácidos graxos.
Não houve significância (P> 0,20) para a interação óleo de soja × monensina para os ácidos graxos avaliados. A monensina não alterou a proporção dos ácidos graxos de cadeias curta (AGCC, C4:0-C10:0), média (AGCM, C12:0-C16:0) ou longa (> que C17:0), mas o óleo de soja reduziu em 43,7% os AGCC e em 49,1% os AGCM, conseqüentemente, aumentando em 55,3% os ácidos graxos de cadeia longa.
A redução da proporção dos AG de cadeias curta e média pode ser decorrente da diminuição de precursores da síntese de novo, acetato e β-hidroxibutirato, resultantes da fermentação ruminal, ou da direta inibição do complexo enzimático envolvido na síntese de novo pela ação dos ácidos graxos de cadeia longa dos óleos vegetais (Palmquist et al., 1993).
Ainda que o óleo de soja e a monensina tenham reduzido a proporção de acetato e de butirato nos AGV totais do líquido ruminal (Eifert et al., 2005a), parece não ter limitado a quantidade de substrato para a síntese de novo. Embora o óleo de soja tenha reduzido os AG de cadeia curta, este efeito foi somente de 9,3% sobre o butirato (C4:0, Tabela 3), o que confirma a idéia de que o butirato é sintetizado de forma independente ao malonil-CoA, sendo a maioria sintetizada a partir da direta incorporação do butirato do rúmen nos lipídios do leite (Chilliard et al., 2000). Além disso, os AG de cadeia média (C12:0 a C16:0) foram reduzidos somente pelo óleo e não foram afetados pela monensina, indicando pouca influência da alteração das proporções dos AGV sobre os ácidos graxos resultantes da síntese de novo. Os efeitos mais pronunciados do óleo de soja foram sobre os ácidos graxos maiores que C6:0, indicando o efeito inibitório dos AG de cadeia longa sobre a Acetil-CoA Carboxilase na síntese de novo.
O óleo de soja reduziu a concentração dos AG de cadeia ímpar (Tabela 2), determinado pelo somatório de C11:0, C13:0, C15:0 e C17:0 (Tabela 3). Os ácidos graxos de cadeia ímpar são originários dos lipídios microbianos sintetizados a partir da utilização de propionato e valerato como primers (Mansbridge & Blake, 1997). Então, a síntese de AG de cadeia ímpar no leite reflete a síntese de AG microbiano, somada à síntese de células microbianas. Os resultados encontrados neste estudo corroboram os obtidos por Eifert et al. (2005a), que constataram que o óleo de soja inibiu a síntese microbiana. A monensina apresentou efeito somente sobre C17:0, mas não no total de ácidos graxos de cadeia ímpar. O processo pelo qual as bactérias regulam sua composição lipídica é pouco conhecido, mas Demeyer & Doreau (1999) indicam que, em altas quantidades de lipídios dietéticos, bactérias e protozoários podem incorporar o ácido linoléico e outros AG em sua estrutura de membrana, inibindo a síntese de novo microbiana, o que justifica a menor proporção de ácidos de cadeia ímpar nas dietas com óleo.
Tabela 2 - Resumo do perfil dos ácidos graxos do leite, em g/100 g de ácidos graxos
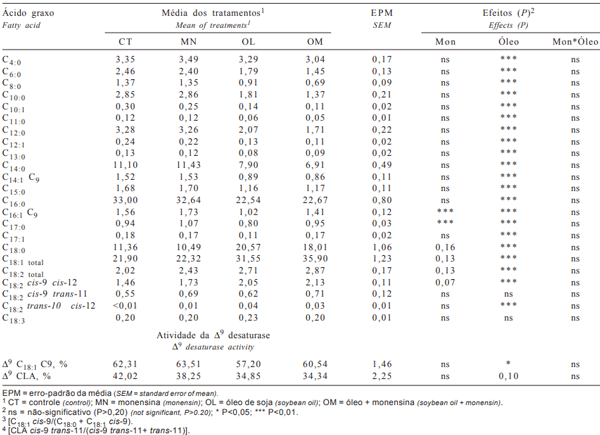
Tabela 3 - Perfil dos ácidos graxos do leite e estimativa da atividade da enzima Δ9 desaturase, em g/100 g de ácidos graxos
Os AG insaturados do leite foram aumentados em 39,2% e os AG saturados reduzidos em 18,1% nas dietas com óleo de soja, o que reflete o perfil de AG da dieta, correspondendo a acréscimos de AG monoinsaturados (AGMI) e poliinsaturados (AGPI), respectivamente, de 39,3 e 24,2%. A monensina proporcionou aumento de 9,0% nos AG insaturados e redução de 4,5% nos AG saturados do leite, com concomitante aumento de 8,8 e 10,7% para AGMI (P=0,14) e AGPI (P=0,06). Sauer et al. (1998) observaram aumentos significativos nestes ácidos graxos com o fornecimento de monensina, sugerindo que este efeito foi ocasionado pela redução na extensão da bio-hidrogenação ruminal.
A concentração do ácido esteárico (C18:0, Tabela 3) elevou na presença do óleo, possivelmente como conseqüência do maior consumo deste ácido graxo e da maior quantidade de AG expostos à bio-hidrogenação. Os valores obtidos nas dietas com óleo são altos em relação às médias encontradas no leite por Chilliard et al. (2000), indicando intensa bio-hidrogenação do óleo de soja. Algumas pesquisas têm comprovado, em ensaios in vitro (Van Nevel & Demeyer, 1995; Fellner et al., 1997) e in vivo (Sauer et al., 1998), que a monensina inibe o último passo da bio-hidrogenação ruminal, diminuindo a concentração do ácido esteárico. Entretanto, neste estudo, embora tenham-se observado valores numericamente menores quando se compararam as dietas CT com MN (8,3%; 11,36 vs 10,49 g/100 g) e OL com OM (14,2%; 20,57 vs 18,01 g/100 g), o efeito da monensina sobre a concentração de C18:0 somente foi significativo na probabilidade de P = 0,16.
Os ácidos graxos insaturados C18:1, C18:2 e C18:3 foram aumentados pelo óleo de soja, enquanto, com monensina, houve elevação das concentrações dos totais C18:1 e C18:2 somente na probabilidade de P = 0,13. Tanto óleo como monensina foram eficientes para elevar as concentrações do ácido linoléico (cis-9 cis-12 C18:2) no leite, embora os valores observados sejam baixos em relação à quantidade de óleo fornecido nas dietas. Chilliard et al. (2000) afirmam que as baixas concentrações de ácido linoléico no leite são decorrentes da baixa eficiência de secreção quando altas quantidades deste ácido são fornecidas na dieta da fase de lactação e da extensão da bio-hidrogenação ruminal. Segundo esses autores, a eficiência de transferência do ácido linoléico no terço inicial é menor que nos terços médio e final da lactação, sugerindo maior preferência de utilização pelo sistema reprodutivo e pelos tecidos de reserva que pela glândula mamária.
Embora os AG do leite e os AG absorvidos no intestino delgado estejam altamente correlacionados, é difícil quantificar a bio-hidrogenação ruminal a partir do perfil de AG do leite, pois os AG da dieta sofrem influência da atividade de desaturação nos intestinos, da seleção de específicos AG pela glândula mamária e dos efeitos dos AG circulantes em função da mobilização corporal. A bio-hidrogenação ruminal é determinada pela proporção dos ácidos oléico, linoléico e linolênico que chegam ao intestino delgado em relação à quantidade ingerida. Duckett et al. (2002) reportaram que a bio-hidrogenação é maior para o ácido linolênico (91%), intermediária para o ácido linoléico (80%) e menor para o ácido oléico (70%).
Diversos autores têm sugerido o acúmulo de transC18:1 no leite como característica de incompleta biohidrogenação dos AG no rúmen (Mansbridge & Blake, 1997). A concentração de trans-C18:1 tem sido aumentada em situações como altas quantidades de lipídios insaturados na dieta (Bateman & Jenkins, 1998), baixo pH ruminal (Kalscheur et al., 1997) e presença de ionóforos (Cant et al., 1997; Sauer et al., 1998). Segundo Bessa et al. (2000), a população microbiana é muito sensível a esses fatores e a maior produção ruminal de isômeros trans-C18:1 nestas situações é um mecanismo de defesa da microbiota, permitindo às bactérias manter a integridade da membrana celular, uma vez que ácidos graxos trans são menos tóxicos que os de configuração cis.
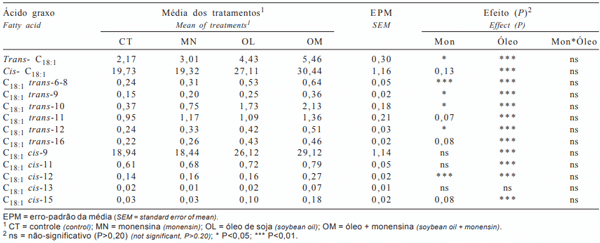
A monensina não influenciou a concentração dos isômeros cis-C18:1 (Tabela 4), enquanto a adição de óleo de soja aumentou sua concentração em 46,9%, possivelmente em razão do maior consumo de ácidos graxos insaturados da dieta, embora o óleo de soja tenha diminuído a atividade da Δ9-desaturase, que converte o C18:0 em cis-9 C18:1 (Δ9 C18:1 C9, Tabela 3). Individualmente, cis-9 C18:1 foi o isômero cis-C18:1 mais abundante e, enquanto a monensina aumentou somente a concentração de cis-12 C18:1, o óleo de soja só não elevou a concentração de cis-15 C18:1.
Tanto o óleo de soja como a monensina aumentaram a concentração dos isômeros trans-C18:1 no leite (Tabela 4). A monensina elevou a concentração dos isômeros transC18:1 no leite, independentemente da presença de óleo, indicando efeito aditivo do óleo de soja e da monensina. Os isômeros trans-9, trans-10, trans-11, trans-12 e trans-16 C18:1 foram aumentados tanto por óleo como por monensina, o que reforça a possibilidade de diferentes rotas de bio-hidrogenação. De acordo com Loor & Herbein (2003), trans-6-8 e trans-9 C18:1 são os principais intermediários durante a isomeração do ácido oléico (cis-9 C18:1) e intermediários menores da biohidrogenação do ácido linolênico. Recentemente, Mosley et al. (2002) verificaram que o ácido oléico não é reduzido diretamente ao esteárico (C18:0), mas parte é isomerada a trans-9, trans-10, trans-11 ou trans-12 C18:1 antes de sua completa saturação.
Tabela 4 - Perfil dos ácidos graxos cis e trans octadecenóicos do leite (g/100 g)
A clássica rota da bio-hidrogenação ruminal do ácido linoléico envolve a formação do CLA cis-9 trans-11 C18:2 e sua redução até trans-11 C18:1 antes da completa saturação até o ácido esteárico, C18:0 (Harfoot & Hazlewood, 1997). Como conseqüência dessa rota metabólica microbiana, 80% do CLA encontrado no leite apresenta a configuração cis-9 trans-11 C18:2, sendo o trans-11 C18:1 o principal isômero intermediário detectado (Bauman & Griinari, 2001). Entretanto, Griinari et al. (1998) sugerem que, dependendo do ambiente ruminal, a rota da bio-hidrogenação ruminal pode ser desviada, promovendo a formação de outros isômeros do CLA e seus trans-C18:1 correspondentes.
A concentração de trans-11 C18:1 foi aumentada pelo óleo de soja e pela monensina (P=0,07). Elevar a concentração deste ácido graxo é de grande importância, uma vez que este AG é o precursor da síntese endógena de CLA cis-9 trans-11 C18:2, que apresenta propriedades anticancerígenas e outras benéficas à saúde humana (Parodi, 1999). Enquanto os tratamentos aumentaram trans-11 C18:1, não foi verificado efeito significativo dos tratamentos sobre a concentração de CLA cis-9 trans-11 C18:2 (Tabela 3), embora as dietas MN, OL e OM tenham aumentado sua concentração em relação à dieta CT em 20,8; 24,5 e 30,2%, respectivamente.
A ausência de resposta estatística para este CLA pode ser efeito do pequeno número de animais amostrados. Apesar do coeficiente de variação de 17,14% verificado neste trabalho, Kelsey et al. (2003), com um número maior de animais, observaram variação de três vezes o conteúdo de CLA entre indivíduos e demonstraram que a base para a variação individual está mais relacionada à passagem pelo rúmen de CLA cis-9 trans-11 C18:2 e de trans-11 C18:1 e à atividade da Δ9-desaturase que a fase de lactação ou o grupo genético dos animais.
Estudos indicam que a principal forma de secreção do CLA cis-9 trans-11 C18:2 no leite é a partir da atividade enzimática da Δ9-desaturase sobre o trans-11 C18:1 (Corl et al., 2001). Neste estudo, estimou-se a atividade desta enzima a partir da relação produto/precursor (CLA cis-9 trans-11 C18:2/[CLA cis-9 trans-11 C18:2+ trans-11 C18:1]), na qual o menor valor indica inibição. O óleo de soja inibiu a atividade desta enzima (Δ9CLA, Tabela 3), o que pode ser interpretado de duas formas: os valores similares da Δ9 CLA e maiores valores de trans-11 C18:1 no leite nas dietas MN em relação à CT indicam que a maior parte do CLA secretado no leite na dieta MN tem origem na Δ9-desaturase. Entretanto, a maior concentração de trans-11 C18:1 e a menor atividade enzimática nas dietas OL e OM sugerem que grande parte do CLA secretado no leite não foi de origem endógena, mas proveniente do CLA cis-9 trans-11 C18:2 gerado no rúmen.
O consumo de AG trans-C18:1 tem promovido redução de HDL e aumento da concentração plasmática de colesterol e LDL e incidência de doenças coronarianas. Portanto, o aumento dos isômeros trans-C18:1 obtido com os tratamentos poderia indicar menor qualidade do leite para a saúde humana. No entanto, segundo Lock & Bauman (2003), a maioria dos estudos não faz distinção entre os efeitos de diferentes isômeros trans-C18:1 e grande parte dos dados que relaciona o consumo destes AG a doenças coronarianas foi obtida com gorduras vegetais parcialmente hidrogenadas, que não proporcionam o mesmo perfil dos isômeros trans-C18:1 do leite.
Nos processos industriais de margarina, há aumento dos isômeros trans-C18:1, que, muitas vezes, atingem 40-50% dos ácidos graxos totais, sobretudo o ácido elaidico (trans-9 C18:1), que tem seus efeitos sobre as doenças coronarianas extrapolados para os demais isômeros trans-C18:1. Lock & Bauman (2003) relataram evidências de que, ao contrário do consumo de gorduras hidrogenadas de origem vegetal, o consumo de trans-11 C18:1 é negativamente correlacionado ao risco de doenças coronarianas. Além disso, os autores demonstraram que a Δ9-desaturase também é ativa no tecido humano, de modo que o consumo de trans-11 C18:1 e o CLA cis-9 trans-11 C18:2 formado, são eficientes por reduzirem o risco de doenças coronarianas e de aterosclerose e a incidência de tumores mamários.
A concentração de trans-10 C18:1 no leite foi aumentada tanto pelo óleo de soja como pela monensina (Tabela 4). Maiores concentrações de trans-10 C18:1 estão associadas a aumentos do CLA trans-10 cis-12 C18:2 e ambos estão associados aos mecanismos que causam a síndrome do baixo teor de gordura do leite (Bauman & Griinari, 2001; Loor & Herbein, 2003). Ainda que somente as dietas com óleo tenham proporcionado aumento na concentração de CLA trans-10 cis-12 C18:2 no leite (Tabela 3), os valores são baixos em relação aos observados com o fornecimento deste CLA na forma de sais de cálcio. Dessa forma, sugere-se que a redução no teor de gordura do leite, quando se utiliza monensina na dieta, pode ser decorrente do aumento de trans-10 C18:1 e CLA trans-10 cis12 C18:2, e não à alteração dos produtos finais da fermentação, como freqüentemente relatado na literatura.
Griinari et al. (1998) promoveram alteração na fermentação ruminal com o uso de dietas ricas em carboidratos e baixos teores de fibra, selecionando uma população microbiana que elevou a concentração de trans-10 C18:1 no leite, refletindo redução do teor de gordura. Em cultura contínua, Jenkins et al. (2003) observaram efeitos de óleo de soja e de monensina sobre a concentração de trans-10 C18:1 e relataram que o comportamento deste AG foi similar ao observado neste trabalho, embora tenham verificado significância para a interação tratamentos × substrato, com maior magnitude na formação de trans-10 C18:1 quando a cevada foi comparada ao milho como fonte de carboidrato.
Do ponto de vista microbiológico, as alterações nas rotas de bio-hidrogenação ruminal podem ser conseqüência do estímulo ou da inibição ao crescimento de determinadas espécies ou grupos de bactérias ruminais por fatores dietéticos – taxa de degradação dos carboidratos, pH ruminal, substrato preferencial – ou do uso de ionóforos.
As bactérias responsáveis pela redução ao trans-11 C18:1, quando na rota mais freqüente da bio-hidrogenação do ácido linoléico, são Butyrivibrio fibrisolvens e Ruminococcus sp., que se comportam como gram-positivas e, portanto, sensíveis à monensina (Nagaraja et al., 1997). Conseqüentemente, a população selecionada pelo ionóforo promoveu a formação de outros isômeros trans-C18:1 e outros isômeros do CLA, justificando os resultados obtidos para os AG trans, sobretudo o trans-10 C18:1. Em estudos com cepas isoladas de Clostridium sp., grampositivas, Bauman & Griinari (2001) relataram a produção de trans-10 C18:1 como principal intermediário da bio-hidrogenação do ácido linoléico. Butyrivibrio fibrisolvens e Ruminococcus sp. necessitam de menor dose de monensina para controle de 50% da população que Clostridium sp. (Nagaraja et al., 1997). Assim, possivelmente a população de Clostridium sp., entre outras presentes no rúmen dos animais avaliados, não foi alterada pela monensina, justificando os aumentos em trans-10 C18:1. Além destas, Kim et al. (2000), verificaram que Megasphaera elsdenii, uma espécie de bactéria que utiliza lactato e resistente à monensina, produz CLA trans-10 cis-12 C18:2 como principal intermediário da bio-hidrogenação do ácido linoléico, o que pode resultar em maior concentração de trans-10 C18:1.
Ainda não foram sintetizados isômeros puros do trans10 C18:1 , o que poderia confirmar os efeitos sobre a síntese de gordura do leite. Entretanto, infusões no abomaso de apenas 3,5 g de CLA trans-10 cis-12 C18:2 promovem redução de 25% no teor da gordura do leite (Baumgard et al., 2001). A magnitude da redução da gordura foi correspondente à inibição de enzimas relacionadas à síntese de novo, à abundância de mRNA que codifica genes envolvidos na absorção e no transporte dos AG, à atividade da Δ9-desaturase e à formação de triglicerídios (Baumgard et al., 2002).
A redução no teor de gordura do leite dos animais deste trabalho (Eifert et al., 2005b) refletiu em aumentos na concentração de trans-10 C18:1 (r=-0,6943; P=0,0001), com efeitos mais pronunciados sobre a proporção dos AG de cadeia média (r = -0,9279; P = 0,0001) e curta (r = -0,8823; P = 0,0004), enquanto foi positivamente correlacionada aos AG de cadeia longa (r = -0,9363; P = 0,0001). Esta alteração da proporção dos AG tem importância sobre a saúde humana, pois os ácidos graxos resultantes da síntese de novo são correlacionados a aumentos do colesterol plasmático e a incidências de doenças coronarianas e cardiovasculares (Parodi, 1999).
Poucas diferenças nos AG foram observadas quando comparados os efeitos dos tratamentos em relação à semana de lactação dos animais. Possivelmente, a ausência de resposta entre a 5a e 15a semanas da lactação decorreu das taxas de ganho de peso e do balanço energético positivo dos animais (Eifert et al., 2005b), o que reduziu o efeito dos ácidos graxos resultantes da mobilização corpórea sobre o perfil da gordura do leite. Esse efeito é ilustrado na Tabela 5, com valores semelhantes para AGCM, C14:0, C16:0, cis-9 C18:1 nas dietas CT e MN.
Tabela 5 - Valores médios observados para os efeitos da interação tratamentos × semana da lactação sobre as concentrações de C14:0, C16:0, C18:1 cis-9 C18:1 e cis-C18:1 totais na gordura do leite (em g/100 g)
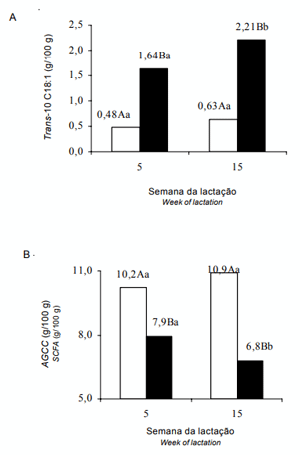
Figura 1 - Interação óleo × semana da lactação das variáveis: trans-10 C18:1 (A), ácidos graxos de cadeia curta - AGCC ( B) do leite; colunas brancas = dietas sem óleo; colunas pretas = dietas com óleo. ‘A’, ‘B’ indica diferença na mesma semana; ‘a’ e ‘b’ indicam diferença dentro do tratamento (Tukey, P< 0,05).
Enquanto as dietas OL e OM propiciaram a produção de leite com maiores valores de cis-9 C18:1 e dos isômeros cis-C18:1 totais em relação às dietas CT e MN, a concentração destes AG foi aumentada da 5a à 15a semanas da lactação com a dieta OL. O reflexo deste aumento pode ser observado na redução dos AGCM, C14:0 e C16:0 nas mesmas semanas da lactação (Tabela 5).
A concentração de trans-10 C18:1 aumentou da 5a à 15a semana da lactação nas dietas com óleo de soja (Figura 1A), sugerindo efeito cumulativo sobre a população microbiana e sua atividade, no sentido de maior produção deste ácido graxo quando óleo de soja foi fornecido na dieta por longo período. Os reflexos da maior proporção de trans-10 C18:1 são a redução da proporção dos ácidos graxos de cadeia curta nas dietas com óleo (Figura 1B) e a menor concentração de AGCM nos tratamentos OL e OM (Tabela 5) da 5a à 15a semana da lactação. Portanto, algum mecanismo de saturação com o trans-10 C18:1, possivelmente com efeito cumulativo, pode ter ocorrido sobre o complexo enzimático na glândula mamária, proporcionando as menores concentrações dos AGCC e AGCM na 15a semana da lactação.
Conclusões
O fornecimento de óleo de soja na dieta altera o perfil de ácidos graxos do leite, aumentando a participação de ácidos graxos insaturados de cadeia longa, enquanto evidências indicam que a monensina aumenta os ácidos graxos insaturados do leite, por inibir a bio-hidrogenação ruminal.
A monensina e o óleo de soja apresentam efeitos aditivos sobre os isômeros trans-C18:1, sobretudo trans-10 C18:1, que pode provocar efeitos cumulativos sobre a população microbiana quando o óleo de soja é fornecido por longo período, resultando em maior inibição do teor de gordura do leite.
Embora os tratamentos não tenham influenciado o conteúdo de CLA cis-9 trans-11 C18:2, tanto óleo de soja como monensina aumentam trans-11 C18:1 no leite, que pode ser convertido em CLA no tecido humano e melhorar a qualidade nutritiva do leite.
Publicado orginalmente na Revista Brasileira de Zootecnia, c., v.35, n.1, p.219-228, 2006. Acesso disponível em: https://www.scielo.br/j/rbz/a/yjTSnchSTrFsXSyRGfvY7kt/?format=pdf&lang=pt