Transferência de imunidade passiva e proteinograma sérico em bezerros das raças Crioula Lageana variedade Mocha e Aberdeen Angus (Red Angus) nos primeiros seis meses de vida
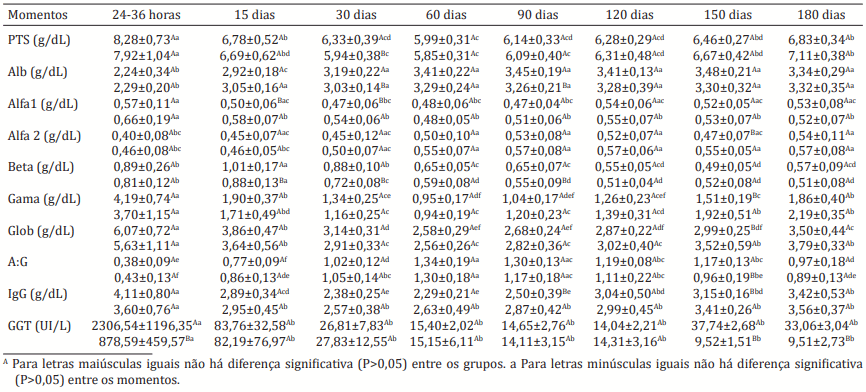
A raça Crioula Lageana apresenta-se como uma alternativa para a pecuária moderna, devido a sua grande variabilidade genética, rusticidade e adaptação às condições ambientais do Planalto Catarinense. Estas características podem influenciar importantes eventos fisiológicos, os quais deveriam ser investigados, pois podem auxiliar no desenvolvimento de futuros programas de melhoramento animal. Sabe-se que a falha da transferência de imunidade passiva (TIP) constitui um importante fator para a elevação dos índices de mortalidade neonatal na espécie bovina. Desta forma, torna-se indispensável à avaliação dos aspectos relacionados à TIP em bezerros da raça Crioula Lageana variedade mocha e a sua comparação com outras raças de corte criadas no Planalto Serrano Catarinense. Com o objetivo de avaliar e comparar a TIP e o proteinograma sérico foram selecionados dois grupos experimentais compostos por 11 bezerros da raça Crioula Lageana variedade mocha e 14 bezerros da raça Aberdeen Angus (Red Angus). Amostras de sangue foram colhidas por meio da venopunção jugular para a mensuração da proteína total sérica (PTS), eletroforese das proteínas séricas em gel de agarose, atividade da gamaglutamiltransferase (GGT) e concentração de imunoglobulinas G (IgG) por meio do método de turvação pelo sulfato de zinco nos períodos entre 24 e 36 horas, 15, 30, 60, 90, 120, 150 e 180 dias de vida. A análise estatística dos dados foi realizada por meio da análise de variância de medidas repetidas (ANOVA), teste de Tukey para a comparação entre médias (P<0,05) e correlação entre as variáveis. Ao analisar os valores de PTS, globulinas, gamaglobulinas, IgG e atividade sérica de GGT ao nascimento (24 a 36 horas) pode se afirmar que houve adequada TIP não havendo diferença significativa entre as raças, exceto para a atividade sérica da GGT. Observaram-se variações fisiológicas na curva de proteínas séricas do nascimento até os 180 dias de idade. Forte correlação positiva foi observada entre os valores de gamaglobulina e IgG (r=0,814 para CL e r=0,877 para AA), PTS (r=0,783 para CL e r=0,947 para AA), globulinas (r=0,945 para CL e r=0,985 para AA), evidenciando que estas variáveis foram bons indicadores da TIP em bezerros ao segundo dia de vida (24–36 horas). A correlação entre gamaglobulinas e atividade da GGT foi fracamente positiva (r=0,251) para bovinos da raça Crioula Lageana variedade mocha e fortemente positiva (r=0,815) para a raça Aberdeen Angus (Red Angus), o qual foi admitido a grande variação nos valores da GGT após 24 horas de vida do bezerro. Não houve diferença na TIP entre as raças Crioula Lageana variedade mocha e Aberdeen Angus (Red Angus), ocorrendo de maneira bem-sucedida em ambas as raças. Pode-se sugerir que a raça Crioula Lageana variedade mocha seja mais tardia na síntese ativa de anticorpos, ressaltando a precocidade dos taurinos evidenciada na raça Aberdeen Angus (Red Angus).
TERMOS DE INDEXAÇÃO: Bovino, imunoglobulinas, imunidade passiva, proteínas séricas.
INTRODUÇÃO
MATERIAL E MÉTODOS
RESULTADOS E DISCUSSÃO
CONCLUSÕES
Amorim R.M. 2002. Valores séricos e hepáticos de elementos minerais, atividade sérica da ceruloplasmina, hemograma, proteinograma e metabolismo oxidativo dos neutrófilos em bezerros da raça Nelore, nascidos de mães com nutrição adequada ou inadequada em cobre e zinco. Tese de Doutorado em Clínica Veterinária, Faculdade de Medicina Veterinária e Zootecnia, Universidade Estadual Paulista, Botucatu, SP. 158p.
Besser T.E. & Gay C.C. 1994. The importance of colostrum to the health of the neonatal calf. Vet. Clin. North Am., Food Anim. Pract. 10(1):107-117.
Bessi R., Pauletti P., d’Arce R.C. & Machado Neto R. 2002. Absorção de anticorpos do colostro em bezerros I. Estudo no intestino delgado proximal. Revta Bras. Zootec. 31(6):2314-2324.
Borges A.S. 1997. Avaliação da eficácia da administração de plasma, por via intravenosa, como tratamento da falência de transferência de imunidade passiva em bezerros da raça Holandesa. Dissertação de Mestrado em Clínica Veterinária, Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo/SP. 84p.
Bouda J. & Jagos P. 1984. Biochemical and hematological reference values in calves and their significance for health control. Acta Vet. Brno 53(3):193-198.
Braun J.P., Rico A.G., Benard P., Thouvenot J.P. & Bonnefis M.J. 1978. Blood and tissue distribution of gamma glutamyl transferase in calves. J. Dairy Sci. 61:596-599.
Braun J.P., Tainturier D., Laugier C., Bénard P., Thouvenot J.P. & Rico A.G. 1982. Early variations of blood plasma gamma-glutamyl transferase in newborn calves: A test of colostrum intake. J. Dairy Sci. 65(11):2178- 2181.
Costa J.N. 2000. Leucograma, metabolismo oxidativo dos neutrófilos, proteinograma e imunoglobulinas de bovinos da raça holandesa (Bos taurus). Influência do desenvolvimento etário e da suplementação com vitamina E (acetato de DL-alfa-tocoferol). Tese de Doutorado em Clínica Veterinária, Faculdade de Medicina Veterinária e Zootecnia, Universidade Estadual Paulista, Botucatu, SP. 209p.
Costa J.N., Peixoto A.P.C., Kohayagawa A. & Souza T.S. 2007. Proteinograma sérico de bezerras da raça Holandesa do nascimento aos 150 dias de idade. Revta Bras. Saúde Prod. Anim. 8(4):267-275.
Costa M.C., Flaiban K.K.M.C., Coneglian M.M., Feitosa F.L.F., Balarin M.R.S. & Lisboa J.A.N. 2008. Transferência de imunidade passiva em bezerros das raças nelore e limousin nos primeiros quarto meses de vida. Pesq. Vet. Bras. 28(9):410-416.
Curi P.R. 1998. Metodologia e análise da pesquisa em ciências biológicas. 2ª ed. Gráfica e Editora Tipomic, Botucatu. 263p.
Delfino J.L., Barbosa V.M., Gondim C.C., Oliveira P.M., Nasciutti N.R., Oliveira R.S.B.R., Tsuruta S.A., Mundim A.V. & Saut J.P.E. 2014. Perfil bioquímico sérico de bezerros Senepol nos primeiros 120 dias de idade. Semina, Ciênc. Agrárias 35(3):1341-1350.
Fagliari J.J., Oliveira E.C., Pegorer M.F., Ferrante Júnior L.C. & Campos Filho E. 1996. Relação entre o nível sérico de gamaglobulinas e as atividades de gama glutamiltransferase, fosfatase alcalina e aspartato aminotransferase de bezerros recém-nascidos. Arq. Bras. Med. Vet. Zootec. 48(2):105-112.
Fagliari J.J., Passipieri M. & Okuda H.T. 1991. Valores de referência das proteínas séricas de bovinos Guzerá em diferentes faixas etárias. Arq. Bras. Med. Vet. Zootec. 43(1):39-60.
Fagliari J.J., Passipieri M., Curi P.R., Ferreira Neto J.M. & Lucas A. 1988. Valores padrões das proteínas séricas de bovinos da raça Guzerá. II. Proteinograma sérico de bezerros recém-nascidos. Ars Vet. 4:225-232.
Fagliari J.J., Santana A.E., Lucas F.A., Campos Filho E. & Curi P.R. 1998. Constituintes sangüíneos de bovinos recém-nascidos das raças Nelore (Bos indicus) e Holandesa (Bos taurus) e de bubalinos (Bubalis bubalus) da raça Murrah. Arq. Bras. Med. Vet. Zootec. 50(3):253-62.
Feitosa F.L.F. 1998. Dinâmica do proteinograma e da atividade da gama glutamiltransferase no soro sangüíneo de bezerros desde o nascimento até um ano de vida e de vacas antes e após o parto, da raça holandesa. Tese de Doutorado em Clínica Veterinária, Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo/ SP. 219p.
Feitosa F.L.F., Birgel E.H., Mirandola R.M.S. & Perri S.H.V. 2001a. Proteinograma sérico de bezerros holandeses do nascimento até um ano de vida. Revta Bras. Ciênc. Vet. 8(2):105-108.
Feitosa F.L.F., Birgel E.H., Mirandola R.M.S. & Perri S.H.V. 2001b. Diagnóstico de falha de transferência de imunidade passiva em bezerros através da determinação de proteína total e de suas frações eletroforéticas, imunoglobulinas G e M e da atividade da gama glutamiltransferase no soro sanguíneo. Ciência Rural 31(2):251-255.
Feitosa F.L.F., Camargo D.G., Yanaka R., Mendes L.C.N., Peiró J.R., Bovino F., Lisboa J.A.N., Perri S.H.V. & Gasparelli E.R.F. 2010. Índices de falha de transferência de imunidade passiva (FTIP) em bezerros holandeses e nelores, às 24 e 48 horas de vida: valores de proteína total, de gamaglobulina, de imunoglobulina G e da atividade sérica de gamaglutamiltransferase, para o diagnóstico de FTIP. Pesq. Vet. Bras. 30(8):696-704.
Garcia R.A.S., Werner O., Barca Junior F.A., Silva L.C., Cunha Filho L.F.C., Lisboa J.A.N., Balarin M.R.S. & Headley S.A. 2012. Evalution of the transfer of passive immunity by the analysis of immunoglobulin and serum proteins of Holstein calves. Semina, Ciênc. Agrárias 33(4):1515-1520.
Halliwell R.E.W. & Gorman N.T. 1989. Veterinary Clinical Immunology. W.B. Saunders, Philadelphia. 548p. Jain N.C. 1993. Essentials of Veterinary Hematology. Lea and Febiger, Philadelphia. 417p.
Kaneko J. J. & Mills R. 1970. Hematological and blood chemical observations in neonatal normal and porphyric calves in early life. Cornell Vet. 60(1):52-60. Kaneko J.J., Harvey J.W. & Bruss M.L. 1997. Clinical Biochemistry of Domestic Animals. 5th ed. Academic Press, San Diego, p.117-138.
Kremers B., Briere R. & Batsakis J.G. 1967. Reflectance densitometry of cellulose acetate protein eletrophoresis. Am. J. Med. Technol. 33(1):28- 34.
Leal M.L.R., Benesi F.J., Lisbôa J.A.N., Coelho C.S. & Mirandola R.M.S. 2003. Proteinograma sérico de bezerras sadias, da raça holandesa, no primeiro mês pós-nascimento. Braz. J. Vet. Res. Anim. Sci. 40(2):138-145.
Logan E.F., Penhale W.J. & Jones R.A. 1972. Changes in the serum immunoglobulins of colostrum fed calves during the first twelve weeks postpartum. Res. Vet. Sci. 14(3):394-397.
Machado Neto R., Packer I.U., Prado G.V.B., Bessi R. & Pauletti P. 2004. Colostral immunoglobulins absorption in Canchim and Nelore calves. Revta Bras. Zootec. 33(6):1544-1547.
Mariante A.S. & Cavalcante N. 2000. Animais do Descobrimento: raças domésticas da história do Brasil. Embrapa-Cenargen, Brasília. 232p.
Martins V.M.V., Veiga T.F., Martins E., Quadros S.A.F., Cardoso C.P. & Ribeiro J.A.R. 2009. Raça Crioula Lageana. O esteio do ontem, o labor do hoje e a oportunidade do amanhã. Editora Associação Brasileira dos Criadores da Raça Crioula Lageana (ABCCL), Lages. 98p.
McGuire T.C. & Adams D.S. 1982. Failure of colostral immunoglobulin transfer to calves: Prevalence and diagnosis. Compend. Contin. Educ. Pract. Vet. 4(1):35-40.
Moraes M.P., Weiblen R., Silva A.M. & Tobias F.L. 1997. Evolução da imunidade passiva em fêmeas bovinas da raça Holandesa. Ciência Rural 27(3):435-40.
Naylor J.M., Kronfeld D.S., Bech-Nielsen S. & Bartholomew R.C. 1977. Plasma total protein measurement for prediction of disease and mortality in calves. J. Am. Vet. Med. Assoc. 171(7):635-638.
Paris T.N.N., Quedraogo G., Rico A.G., Bezzile P. & Braun J.P. 1992. Interêt de la gamma-glutamyl transférase sérique et de la protidêmie totale pour le controle de la prise colostrale chez les veaux nouveaunés. Rec. Méd. Vet. 168:43-47.
Pauletti P., Machado Neto R., Packer I.U., d’Arce R.D. & Bessi R. 2003. Quality of colostral passive immunity and pattern of serum protein fluctuation in newborn calves. Scientia Agricola 60(3):453-456.
Pfeiffer N.E., McGuire T.C., Bendel R.B. & Weikel J.M. 1977. Quantitation of bovine immunoglobulins: comparison of single radial immunodiffusion, zinc sulfate turbidity, serum electrophoresis, and refratometer methods. Am. J. Vet. Res. 38(5):693-698.
Piazza W.F. 1983. Santa Catarina: Sua história. 19ª ed. UFSC Lunardelli, Florianópolis. 750p.
Pires Júnior J.B., Buonora C.R.A.R., Afonso A.B., Dantas F.R., Pereira A.L.L., Vieira A.C.S. & Mendonça C.L. 2013. Transferência de imunidade passiva em bezerros oriundos de partos distócicos obtidos por cesariana. Rev. Bras. Med. Vet. 35(2):111-116.
Radostits O.M., Gay C.C., Blood D.C. & Hinchcliff K.W. 2002. Clínica Veterinária: Um tratado de doenças dos bovinos, ovinos, suínos, caprinos e equinos. 9ª ed. Guanabara Koogan, Rio de Janeiro, p.102-136.
Ribeiro M.F.B., Salcedo J.H.P., Belém P.A.D. & Faria J.E. 1983. Hipogamaglobulinemia em bezerros. Arq. Bras. Vet. Zootec. 35(4):537-546.
Rocha T.G., Niciti R.P., Sampaio A.A.M. & Fagliari J.J. 2012. Passive immunity transfer and serum constituents of crossbred calves. Pesq. Vet. Bras. 32(6):515-522.
Rogers R.W., Martin S.W. & Meek A.H. 1985. Reproductive efficiency and calf survival in Ontario bovine cow-calf herds: A cross sectional mail survey. Can. J. Comp. Med. 49(1):27-33.
Selim S.A., Smith B.P. & Cullor J.S. 1995. Serum immunoglobulins in calves: their effects and two easy reliable means of measurement. Vet. Med. 90:387-404.
Spritze A. 2001. Uso de marcadores moleculares RAPD na caracterização genética da raça bovina Crioulo Lageano. Dissertação de Mestrado em Ciências Agrárias, Universidade de Brasília, Brasília, DF. 67p.
Spritze A., Egito A.A., Mariante A.S. & McManus C. 2003. Caracterização genética da raça bovina Crioulo Lageano por marcadores moleculares RAPD. Pesq. Agropec. Bras. 38(10):1157-1164.
Szasz G. 1969. A kinetic photometric method for serum gamma-glutamyl transpeptidase. Clin. Chem. 15(2):124-136.
Teixeira W.T., Fonteque G.V., Ramos A.F., Mariante A.S., Egito A.A., Martins V.M.V., Saito M.E. & Fonteque J.H. 2012. Transfer of passive immunity and serum proteinogram in the first six months of life of Criollo Lageano and Black and White Holstein calves. Pesq. Vet. Bras. 32(10):980- 986.
Tennant B. & Guerra M.R. 1969. Neonatal alterations in serum gamaglobulin levels of Jersey and Holstein-Friesian calves. Am. J. Vet. Res. 51(2):345-354.
Tizard I.R. 2002. Imunologia Veterinária: uma introdução. 6ª ed. Roca, São Paulo. 120p.
Veiga T.F., Quadros S.A.F., Martins E. & Improta C.T.R. 2009. Raça Crioula Lageana: percepções em relação às possibilidades de sua exploração na região do planalto catarinense. Revta Bras. Agroec. 4(1):29-38.
Vettorato E.D., Costa M.C., Flaiban K.K.M.C., Vidotto O., Balarin M.R.S. & Lisbôa J.A.N. 2012. Variação de proteínas séricas em bezerros das raças nelore e holandesa do nascimento até os seis meses de vida. Semina, Ciênc. Agrárias 33(2):3181-3190.
Wittum T.E. & Perino L.J. 1995. Passive immune status at postpartum hour 24 and long-term health and performance of calves. Am. J. Vet. Res. 56(9):1149-1154.
Zanetti M.A., Lucci C.S. & Lobo R.B. 1982. Duração do período de absorção de imunoglobulina do colostro por bezerros recém-nascidos. Revta Soc. Bras. Zootec.11(4):612-621.
Zanker I.A., Hammon H.M. & Blum J.W. 2001. Activities of ɣ-glutamyltransferase, alkaline phosphatase and aspartate-aminotransferase in colostrum, milk and blood plasma of calves fed first colostrum at 0-2, 6-7, 12-13 and 24-25 h after birth. J. Vet. Med. A, Physiol. Pathol. Clin. Med. 48(3):179-185.
