RESUMO.
Este trabalho foi realizado com o objetivo de avaliar o efeito de suplementos múltiplos contendo palma forrageira enriquecido com uréia (0, 1, 2 e 3% da matéria seca - MS) em substituição a um suplemento tradicional (testemunha) sobre parâmetros ruminais e síntese de proteína microbiana em Cinco novilhos mestiços não ruminais (½ Dutch x Zebu), com peso corporal inicial de 240 ± 22,1 kg foram utilizados em um quadrado latino 5 × 5. Foi observado efeito quadrático para consumo de MS e retenção de N, com valores máximos de 6,97 kg d -1 e 50,9 gd -1 com a inclusão de 1,8 e 2,1% de uréia, respectivamente. Concentrações máximas de 16,2; 23,2 e 24,3 mg dL-1 de N-NH3 foram registrados em animais alimentados com palma forrageira enriquecida com 1,2 de uréia a 3%. Há um efeito quadrático sobre o pH ruminal, com valor de 6,45 a 4, 08 horas após alimentação. A síntese proteica microbiana de 103 g de proteína bruta kg -1 de nutrientes digestíveis totais foi obtida com a inclusão de 1,6% de uréia. Recomenda-se o uso de suplementos múltiplos contendo palma forrageira enriquecido com 1,6 a 1,8% de uréia em substituição ao suplemento tradicional para bovinos.
Palavras-chave: ingestão, pH ruminal, suplementação, ácidos graxos voláteis.
Introdução
O estabelecimento de um manejo eficiente das fêmeas de recria é a base de qualquer sistema de produção leiteira, com participação significativa nos custos de produção (Queiroz, Berchielli, Signoretti, Ribeiro, & Morais, 2012). As pastagens tropicais são a principal fonte de nutrientes para esta categoria animal devido ao baixo custo e alta comodidade.
No entanto, animais criados em pastagens tropicais podem apresentar deficiências múltiplas de nutrientes, principalmente durante o período de dormência das gramíneas, induzidas pelo déficit hídrico registrado na estação seca. melhorar a ingestão de nutrientes, aumentar a concentração de energia da dieta, potencializar os precursores bioquímicos e, assim, promover maior precocidade e desempenho animal (Zervoudakis et al., 2008; Villela et al., 2011).
A proteína, seguida da energia, é o nutriente mais requerido pelos ruminantes. O aproveitamento da forragem pelos bovinos, principalmente quanto aos compostos fibrosos, está diretamente relacionado à atividade microbiana ruminal, que depende do teor de compostos nitrogenados no rúmen (Costa et al., 2011). Em geral, 50 a 70% do nitrogênio microbiano pode ser derivado da amônia ruminal, e o restante, peptídeos e aminoácidos da dieta (Santos et al., 2010).
A palma forrageira, antes considerada uma alternativa alimentar, tornou-se importante nos sistemas de produção leiteira, devido à elevada produção de matéria seca por unidade de área. Além disso, é uma excelente fonte de energia proveniente dos carboidratos não fibrosos e nutrientes digestíveis totais (Ferreira, Bispo, Rocha, Urbano, & Costa, 2012). Assim, os 'suplementos múltiplos' são utilizados em sistemas a pasto para manejar déficits de forragem, podendo ser compostos por uma mistura controladora (ex. uréia + mistura mineral) para regular o consumo dos animais (Valente et al., 2011 ). Diante disso, levantamos a hipótese de que a palma forrageira enriquecida com uréia, na forma de 'suplementos múltiplos', poderia substituir os suplementos tradicionais compostos de milho, farelo de trigo e farelo de soja para bovinos.
Este estudo foi realizado com o objetivo de avaliar o efeito dos 'suplementos múltiplos' contendo palma forrageira enriquecido com uréia, em substituição a um suplemento tradicional sobre a fermentação ruminal e síntese de proteína microbiana em novilhos mestiços.
Materiais e métodos
Este estudo foi realizado no Departamento de Zootecnia da Universidade Federal Rural de Pernambuco, localizada em Recife, Pernambuco, Brasil.
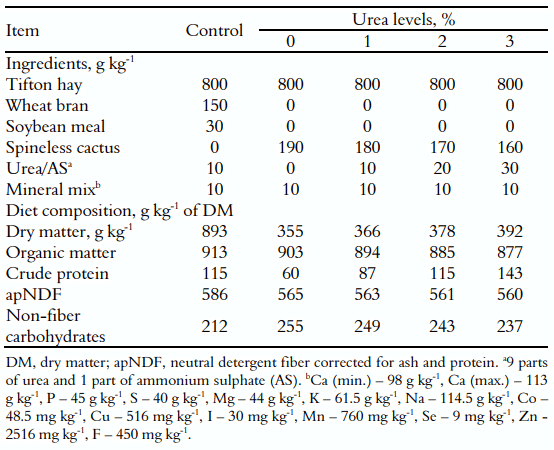
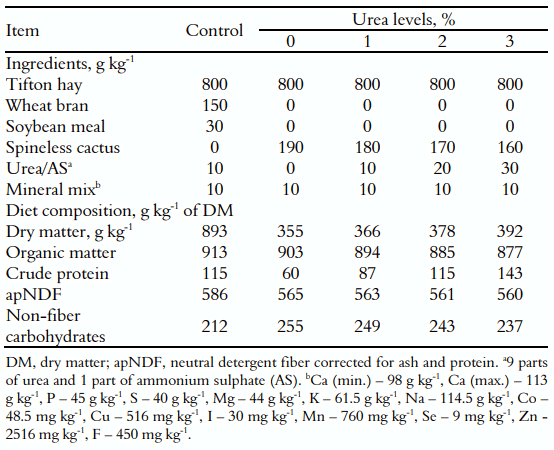
As dietas foram formuladas para atender às exigências do gado leiteiro de acordo com o National Research Council (NRC, 2001). A relação forragem:concentrado foi de 80:20 com base na matéria seca (MS) com feno de Tifton85 (Cynodon spp.) como forragem, que foi utilizado para simular a pastagem. As rações consistiram de quatro níveis de inclusão de uréia/sulfato de amônio (0, 1, 2 e 3% sobre MS) e um tratamento controle composto por um tradicional 'suplemento múltiplo' (composto por farelo de trigo, farelo de soja, uréia e minerais). A uréia + sulfato de amônio foi utilizada para corrigir a proteína da palma forrageira (SC). A mistura dos ingredientes foi realizada manualmente nos comedouros, destacando-se que a mucilagem SC permitiu uma agregação uniforme da uréia.As proporções dos ingredientes e a composição química das dietas são apresentadas na Tabela 1.
O manejo e cuidado dos animais foram realizados pelas diretrizes e recomendações do Comitê de Ética em Estudos Animais da Universidade Federal Rural de Pernambuco (Licença nº009/2015), Recife, Brasil.
Tabela 1. Proporção dos ingredientes e composição química das dietas experimentais (g kg -1 com base na matéria seca).
Cinco novilhos fistulados no rúmen (½ Holandês x Zebu) com peso corporal médio inicial (PC) de 240 ± 22,1 kg foram mantidos em baias individuais e utilizados em delineamento quadrado latino 5 x 5. O experimento durou 80 dias, com cinco períodos consecutivos de 16 dias, e foi dividido em adaptação de 7 dias (Menezes et al., 2011) e amostragem de 9 dias.
A ração foi fornecida em duas refeições diárias às 6:00 e 18:00 horas e as sobras foram pesadas diariamente para obtenção de no máximo 5 a 10% de sobras. As sobras foram amostradas, colocadas em sacos rotulados e armazenadas no freezer para posterior análise. Os ingredientes das dietas foram amostrados semanalmente. Ao final de cada período experimental, as amostras de ração e as sobras foram descongeladas e submetidas à pré-secagem a 60°C por 72h e, posteriormente, moídas em moinho de facas tipo Wiley com malha de 1 mm. Amostras compostas foram salvas para cada animal com base no peso seco de cada período.
Durante três dias (9º ao 11º dia) em cada período experimental, após o fornecimento da dieta matinal, foi realizada coleta total de urina (24 h) e o pH medido a cada seis horas para garantir que fosse mantido abaixo de 3,0. Funil coletores foram acoplados a novilhos para coleta de amostras de urina, que foi realizada em um recipiente contendo 500 mL de ácido sulfúrico a 10% (McSweeney & Denman, 2007). Ao final de cada período de coleta, foram determinados o peso e o volume total de urina, e o teor de N total foi determinado pelo método de Kjeldahl citado por Detmann et al. (2012) (método INCT-CA N004/1). Essas amostras foram armazenadas a -15°C para posterior análise de uréia, creatinina, all an to in e ácido úrico.
Quatro horas após o consumo da dieta, no 11º dia de cada período experimental, foi coletado sangue por punção da veia jugular em tubo de ensaio contendo gel de separação com ativador de coagulante (SST II Advance, BD Vacutainer, Brasil). Essas amostras foram armazenadas a -15°C até posterior análise da uréia.
Amostras de rações, sobras e fezes foram analisadas quanto à matéria seca (MS; método 934.01), matéria orgânica (MO; método 930.05), proteína bruta (PB; método 968.06) e extrato etéreo (EE; método 920.39) de acordo com a Associação of Official Analytical Chemists (AOAC, 2000). Para a análise da FDN foi utilizada uma alfa amilase termoestável, sem uso de sulfito de sódio, e corrigida para cinzas residuais (Mertens, 2002). A FDN também foi ajustada para os teores de compostos nitrogenados pelo método descrito por Licitra, Hernandez e Van Soest (1996).
A quantificação dos teores de carboidratos não fibrosos (CNF) foi realizada de acordo com Detmann e Valadares (2010) da seguinte forma: CNF=1000-[(CP - CPu + U) + FDNap + EE + cinzas]; onde CPu = teor de CP da uréia, U = teor de uréia e apNDF = FDN corrigido para cinzas residuais e proteínas. Os demais termos foram previamente definidos, sendo todos expressos em g kg -1 MS. Os nutrientes digestíveis totais (NDT) foram determinados de acordo com Weiss (1999): NDT = PB d + CNF d + FDN d + EE d × 2,25 (subscrito significa digestível).
A estimativa do balanço de N foi obtida subtraindo-se os valores de excreção fecal e urinária do N ingerido. Determinar a eficiência de utilização do composto de N da dieta; foram utilizados os seguintes indicadores: N-uréia no plasma, excreção urinária de N-uréia e balanço de N. O N-uréia do plasma e da urina foi estimado pelo fator 0,466, segundo Cruz et al. (2006).
As perdas urinárias endógenas foram estimadas pela regressão entre a excreção urinária de nitrogênio (Y) e a ingestão de nitrogênio (X), representadas pelo intercepto e pelo coeficiente de regressão. As perdas endógenas totais, incluindo fecal e urinária, foram estimadas pela regressão do balanço de nitrogênio (Y) e ingestão de nitrogênio (X), expressos em g por kg 0,75 , representados pelo intercepto de regressão.
A análise dos derivados purínicos (DP), alantoína e ácido úrico foi realizada pelo método colorimétrico de Fujihara, ∅rskov, Reeds e Kyle (1987), descrito por Chen e Gomes (1992). A excreção de DP foi calculada multiplicando-se o volume de urina, estimado em 24h, pela concentração de DP das amostras spot de urina. As purinas absorvidas (Y, mmol d -1 ) foram calculadas a partir da excreção de DP (X, mmol d -1 ), utilizando a equação Y = 0,85X + 0,385 PV 0,75 . Onde, 0,85 é a recuperação de purinas absorvidas como DP e 0,385 BW 0,75 é a contribuição endógena para a excreção de purina (Verbic, Chen, MacLeod & Ǿrkov, 1990).
A produção de compostos nitrogenados microbianos foi calculada pela equação Nmic (g Nd -1 ) = (70 x PD absorvida)/(0,83 x 0,116 x 1000). Onde N é o teor de purinas (mg Nmmol -1 ), 0,83 representa a digestibilidade intestinal das purinas e 0,116 é a razão média N-purinas:Ntotal nas bactérias isoladas do rúmen (Chen & Gomes, 1992).
O fluido ruminal foi coletado de novilhos em três dias consecutivos (11º ao 13º dia) às 6h00 e 8h00 em dias consecutivos, e 10h00 e 12h00 no mesmo dia. Um total de 250 mL foi coletado das localizações dorsal anterior, ventral anterior, ventral média, dorsal posterior e ventral posterior dentro do rúmen usando uma seringa de 50 mL aparafusada a um tubo de aço inoxidável que termina com uma sonda coberta por uma malha metálica fina . O fluido ruminal foi acidificado a um pH de 2 usando uma alíquota de 40 mL fixada com 1 mL de H 2 SO 4(1:1) e congelado (-20°C) para posterior análise da concentração de N-NH3. Uma segunda alíquota (47 mL) foi fixada com 3 mL de ácido clorídrico (6 N) e congelada para posterior avaliação da concentração de VFA's. A análise de VFA foi realizada usando um cromatógrafo a gás equipado com um detector de ionização de chama e auto-injetor e equipado com uma coluna GP (30 m × 0,250 mm, 0,25 μm; Chromosorb WAW).
As variáveis estudadas foram analisadas por procedimentos estatísticos utilizando o procedimento PROC MIXED do SAS (Statistical Analysis System, versão 9.1.), adotando-se 0,10 como nível crítico de probabilidade para um erro tipo I, conforme o seguinte modelo:
Y ijk = μ + Ti + A j + P k + ε ijk
onde Y ijk = variável dependente medida no animal j que foi submetido ao tratamento i no período k; μ = média geral, T i = efeito fixo do tratamento i, A j = efeito aleatório do animal j, P k = efeito aleatório do período k, e ε ijk = erro aleatório não observado assumindo distribuição anormal.
O teste de Dunnett foi usado para comparar cada grupo de tratamento (níveis de ureia) com a média do grupo de controle (caracterizado por um tradicional 'suplemento múltiplo'). As comparações entre os níveis de uréia nos suplementos foram realizadas pela decomposição da soma dos quadrados em contrastes ortogonais para efeitos lineares, quadráticos e cúbicos quando apropriado (p < 0,10), com posterior ajuste das equações de regressão. Os contrastes foram: I – Todos os tratamentos versus controle; II – palma forrageira sem uréia versus palma forrageira com uréia; III – Efeito linear nos teores de uréia; IV – Efeito quadrático nos níveis de uréia. O pH ruminal, nitrogênio amoniacal ruminal (RAN) e ácidos graxos voláteis (AGVs) foram analisados como efeitos de medidas repetidas ao longo do tempo, de acordo com o seguinte modelo:
Y ijk = μ + Ti + (t*p) ik + σ ij + P k + ε ijk ,
onde Yijk = observaçãoijk; μ média geral; Ti = efeito do tratamento i; p k = efeito do período k; (t*p) ik = efeito da interação entre o tratamento i e o período k; σ ij = erro aleatório com média 0 e variância σ²σ, a variância entre animais (sujeitos) dentro do tratamento e é igual à covariância entre medidas repetidas dentro dos animais; ε ijk = erro aleatório com média 0 e variância σ², a variância entre as medidas dentro dos animais (Kaps & Lamberson, 2004).
Resultados e discussão
O consumo de MS apresentou comportamento quadrático (p < 0,10) com valor máximo de 6,97 kgd -1 estimado com palma forrageira enriquecida com 1,8% de uréia (Tabela 2). O aumento dos níveis de uréia promoveu aumento linear da ingestão de N (p < 0,001) (Tabela 2). Houve efeito quadrático (p < 0,10) no balanço de N, com retenção máxima de 50,9 g Nd -1 com a inclusão de 2,1% de uréia (Tabela 3). Nas dietas com palma forrageira sem ureia e com 1% de ureia, foi encontrado o menor (p < 0,001) teor de NUP (Tabela 2).
O comportamento observado para o consumo de nitrogênio nos animais alimentados com palma forrageira sem uréia e os alimentados com palma forrageira enriquecida com 1% de uréia foi devido tanto ao consumo de matéria seca dos animais quanto ao teor de proteína da dieta. Segundo Costa et al. (2011), a associação entre a ingestão de nitrogênio e o balanço dos compostos nitrogenados não deve ser interpretada diretamente, pois o organismo do animal não consegue absorver todo o nitrogênio existente no suplemento. Além do aporte proteico no intestino, outros mecanismos estão envolvidos no balanço dos compostos nitrogenados, em especial, dietas com fontes proteicas degradáveis no rúmen promovendo seu aumento. O excesso de amônia ruminal pode gerar perdas tanto de energia quanto de nitrogênio, aumentando o gasto energético do animal, pois a conversão de amônia em uréia custa 12 kcalg-1 de nitrogênio ao animal (Van Soest, 1994).
De acordo com Hoffman et al. (2001), existe uma relação linear entre a quantidade de nitrogênio ingerida e sua excreção, inclusive nas fezes e na urina. Os resultados obtidos neste estudo estão de acordo com essa afirmação, pois o tratamento sem uréia promoveu menor excreção de nitrogênio via urina, enquanto os demais tratamentos promoveram excreção linear de nitrogênio urinário, conforme a inclusão de uréia na dieta.
As menores concentrações de NUP nos animais alimentados com palma forrageira sem uréia e naqueles que receberam palma forrageira enriquecida com 1% de uréia podem ser explicadas porque a uréia transferida do sangue para o epitélio ruminal é rapidamente degradada por bactérias ureolíticas aderidas ao epitélio ruminal. Assim, há uma diferença de potencial entre o rúmen e a corrente sanguínea, garantindo um gradiente favorável à transferência, muitas vezes de forma passiva (Van Soest, 1994). Segundo Chizzotti et al. (2006), o nitrogênio uréico plasmático apresenta alta correlação com os teores de proteína da dieta, justificando assim o aumento linear dos níveis de uréia de inclusão.
Os compostos nitrogenados metabólicos fecais (FMN) corresponderam ao consumo de 3,56 g Nkg -1 MS e foram estimados a partir da equação Ŷ = -3,5581 + 0,9381X (r² = 0,97), sendo Y = nitrogênio digestível e X = nitrogênio ingerido. A estimativa do nitrogênio uréico endógeno (EUN) de 0,265 g Nkg -1 de peso metabólico foi obtida pela excreção urinária de nitrogênio total (Y) e ingestão de nitrogênio (X) (Ŷ = -0,2652 + 0,4743X, r 2 = 0,43). As perdas endógenas (fecal e urinária) de nitrogênio (0,277 g Nkg -1 de peso metabólico) foram obtidas por regressão entre balanço de N (Y) e ingestão de N (X) (Ŷ = -0,2768 + 0,6108X, r 2 = 0,72).
Tabela 2. Consumo de matéria seca (CMS) e balanço de N (NB) em novilhos mestiços alimentados com suplementos múltiplos contendo palma forrageira enriquecida com uréia.
O nitrogênio metabólico fecal (FMN) e as perdas urinárias endógenas estimadas neste estudo (3,56 g Nkg -1 ingestão de MS e 0,265 g N por kg 0,75 ) foram diferentes dos relatados por Valadares, Gonçalves, Rodrigues, Valadares e Silva (1997) em novilhos Zebu (consumo de 5,98g Nkg -1 MS e 0,220 g N por kg 0,75 ) As perdas endógenas totais de nitrogênio (0,277 g N por kg 0,75 ) foram semelhantes às encontradas por Valadares et al. (1997) de 0,246 g de N por kg 0,75. Segundo Benedeti et al. (2014), a excreção urinária de N aumenta linearmente com a inclusão de uréia nas dietas. Portanto, um aumento na perda urinária de N com níveis crescentes de uréia nas dietas é devido à rápida hidrólise ruminal, resultando em escape de nitrogênio amoniacal ruminal do rúmen. Os efeitos do balanço de N, evidenciados pelas perdas totais, refletiram positivamente com a inclusão de uréia nas rações.
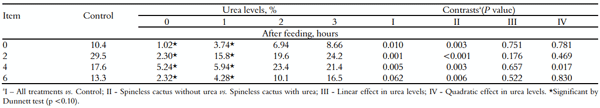
Houve efeito quadrático (p < 0,001) sobre o pH ruminal ao longo do tempo, com pH mínimo de 6,45 às 4,08 horas após a alimentação (Tabela 3). Além disso, o teor de RAN apresentou um comportamento quadrático (p < 0,10) ao longo do tempo quando comparado o tratamento controle e os níveis de uréia (1, 2 e 3% MS). O conteúdo máximo de RAN de 25,0; 11.8; 23,2 e 24,3 mg dL -1 foi estimado em 2,94; 2,76; 3,20 e 3,40 horas após a alimentação, respectivamente (Tabela 4).
A falta de resposta ao pH ruminal neste estudo demonstrou melhora no equilíbrio do ambiente ruminal com a presença da palma forrageira nos 'suplementos múltiplos'. O pH ruminal médio de 6,57 (Tabela 4) manteve-se em níveis considerados adequados (6,2–7,0) por Hoover (1986), o que é justificado pela presença de fibra efetiva do feno, mucilagem da palma forrageira e inclusão de uréia no dietas.
A fibra e a mucilagem estimulam a salivação, evitando a queda do pH, e a uréia promove a alcalinização no rúmen devido ao N-amoníaco produzido. Essa produção de amônia foi decorrente do aumento da ingestão de nitrogênio não protéico (NPN), proveniente da uréia, o que provavelmente levou a um maior teor de RAN (variando de 11,8 a 24,3 mg dL -1 ) nos bovinos alimentados com 'suplementos múltiplos' contendo 1 a 3% de ureia. Este teor de RAN corrobora com a quantidade (10 a 20 mg dL -1 ) sugerida por Leng (1990) para crescimento microbiano máximo.
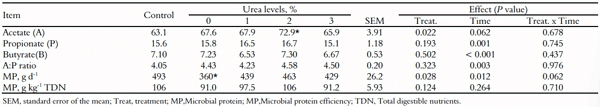
A concentração máxima (p < 0,10) de acetato de 70,9 mmol mL -1 foi estimada com palma forrageira enriquecida com 1,5% de uréia (Tabela 5). Propionato e butirato ao longo do tempo apresentaram concentração máxima (p < 0,001) de 17,5 e 7,8 mmol mL -1 às 4,5 e 4,9 horas após a alimentação, respectivamente (Tabela 5). Os valores máximos (p < 0,10) para proteína microbiana (463 g CPd -1 ) e eficiência microbiana (103 g CPkg -1 NDT) foram estimados com níveis de uréia de 1,9 e 1,6%, respectivamente (Tabela 5).
Tabela 3. pH ruminal de bovinos alimentados com suplementos múltiplos.
Tabela 4. Nitrogênio amoniacal ruminal (RAN, mg dL -1 ) em função dos tempos de coleta.
Tabela 5. Ácidos graxos voláteis (mmol mL -1 ) em novilhos mestiços alimentados com suplementos múltiplos contendo palma forrageira enriquecido com uréia.
Consequentemente, a maior concentração (70,9 mmolmL -1 ) de ácido acético foi estimada com palma forrageira enriquecida com 1,5% de uréia. Apesar do maior teor de carboidratos solúveis e NPN das dietas, não houve efeito sobre a concentração de propionato e butirato. Esse resultado provavelmente é explicado pelo fato de que os carboidratos solúveis e a pectina presentes na palma forrageira são preferencialmente fermentados pelos microrganismos, produzindo tanto acetato quanto etanol (Valadares Filho & Pina, 2011).
Os animais suplementados com palma forrageira sem uréia apresentaram menor síntese de proteína microbiana. Este resultado provavelmente está relacionado ao menor consumo de MS e restrição do fornecimento de N, gerando um perfil fermentativo inadequado. Possivelmente, a composição nutricional da dieta levou a uma maior relação energia:proteína, que por sua vez pode ter limitado a taxa de degradação da proteína, causando uma redução na síntese e eficiência da proteína microbiana. Por outro lado, a redução na proteína microbiana e na eficiência microbiana utilizando palma forrageira acrescida de 2% de uréia foi relacionada ao excesso de RAN e à falta de energia disponível no rúmen. O ingrediente mais energético das dietas, a palma forrageira, teve seu teor reduzido com a adição de uréia.
Nocek e Russell (1988) relataram que a eficiência do crescimento microbiano depende da partição de energia na mantença e no crescimento e está inversamente relacionada ao tempo de residência dos microrganismos no ambiente ruminal. Nesse sentido, quanto mais rápida a passagem dos microrganismos pelo trato gastrointestinal, menos energia é gasta para manutenção, promovendo assim uma maior eficiência da síntese microbiana. Nas condições tropicais, as análises de um conjunto de dados brasileiros sugeriram que 120 g kg -1 de NDT seria mais confiável (Valadares et al., 2010). No presente estudo, o valor médio da eficiência da proteína microbiana (103 g PB kg -1 NDT) obtido por 'suplementos múltiplos' contendo palma forrageira mais 1,6% de uréia foi ligeiramente próximo a este dado.
Conclusão
Assim, 'suplementos múltiplos' contendo palma forrageira enriquecido com 1,6 a 1,8% de uréia em substituição a um suplemento tradicional é recomendado para bovinos por melhorar a ingestão de matéria seca e a fermentação ruminal, favorecendo a síntese de proteína microbiana.