Introdução
A Doença de Gumboro ou Doença Infecciosa da Bursa de Fabrício – DIB caracteriza-se por uma enfermidade aguda, altamente contagiosa, que acomete frangos de corte na fase de crescimento, nos quais a bolsa cloacal, órgão responsável pelo desenvolvimento do sistema imunológico das aves, é primariamente envolvida (ALLAN et al., 1972). É uma doença cosmopolita, que tem causado problemas sanitários na avicultura industrial, pela imunossupressão causada por esta doença, que resulta em perdas econômicas consideráveis (SAPATS et al., 2006).
O controle da doença se resume em medidas de biosseguridade, aliadas aos programas de vacinação, que podem ser definidos de duas formas: o primeiro consiste na proteção de aves jovens por meio da imunidade passiva, em que a vacinação das reprodutoras, no período pré-postura com vacinas oleosas, tem o objetivo de induzir altos e uniformes títulos de anticorpos maternos, protegendo os pintos contra infecções precoces no campo. O segundo se baseia na vacinação da progênie com vacinas vivas, com o intuito de induzir adequada resposta imune ativa contra o vírus de campo de alta virulência durante a fase de criação dos frangos (NISHIZAWA et al., 2007).
Após o registro da presença de cepas mais virulentas da DIB em todo o mundo, por volta do ano de 2002, as vacinas mais leves aplicadas nos pintinhos têm se mostrado menos eficazes. Entretanto, os altos níveis de anticorpos maternos decorrentes dos programas vacinais utilizados nas matrizes podem interferir com a eficácia das vacinações na progênie com vacinas intermediárias. Por outro lado, vacinas atenuadas com menor número de passagens, ou seja, mais agressivas, que por serem imunossupressoras, podem, segundo Giambrone e Closser (1990), resultar em lesões na bolsa cloacal das aves vacinadas.
As reações observadas após a vacinação para outros agentes infecciosos são, geralmente, mais severas em aves acometidas pela DIB, além dos efeitos secundários que afetam negativamente os parâmetros produtivos dos lotes de frangos. Para evitar essa situação, substâncias adjuvantes ou imunomoduladoras, que possam ser facilmente administradas com a vacina, sem causar efeitos adversos e ampliar a resposta imune, estão sendo estudadas (GIAMBRONE; CLOSSER, 1990).
Há duas décadas, o aminoácido arginina (Arg) emergiu como importante modulador da imunidade e de processos fisiológicos. Este aminoácido, além de constituir peptídeos e proteínas em todos os organismos vivos, é precursor de muitos compostos nitrogenados que possuem importantes funções fisiológicas (TAYADE et al., 2006). Duas rotas do metabolismo da Arg são identificadas e conhecidas por terem efeitos imunomodulatórios diretos. A primeira refere-se à Arg que se converte à ornitina e gera poliaminas, as quais possuem papel-chave na divisão celular, síntese de DNA e regulação do ciclo celular. A segunda corresponde à síntese do óxido nítrico (NO), um radical livre altamente reativo, permeável às células e membranas que participa de vários processos celulares, incluindo a neurotransmissão e a imunidade. Altas concentrações de NO podem ser induzidas por uma variedade de estímulos inflamatórios como os lipopolissacarídeos de bactérias e citocinas. Essa rota é essencial para a atividade citotóxica de macrófagos (LE FLOC’H et al., 2004).
Hunphrey e Kirk. (2005), ainda, relataram que a captação de Arg e lisina (Lys), pelo fígado e pela bolsa cloacal durante a resposta do sistema imune, é intensificada para a síntese de proteínas de resposta da fase aguda, indicando que os componentes dietéticos são essenciais para o desenvolvimento dos órgãos linfoides e para uma resposta imune efetiva.
A Arg é considerada um aminoácido essencial para aves, principalmente na fase inicial, pelo fato de que o ciclo bioquímico da ureia não ser funcional em aves (AUSTIC; NESHEIM, 1971), as quais não podem sintetizar Arg ‘de novo’ e por isso são dependentes do fornecimento deste aminoácido nas dietas. Entre as espécies animais estudadas, as aves têm a mais alta exigência de Arg (BALL et al., 2007), que se deve, além da falta de síntese endógena, a alta taxa de deposição proteica pelo rápido crescimento das atuais linhagens de corte, além da interação metabólica entre Lys e Arg, em função da relação antagônica existente entre esses aminoácidos.
A finalidade deste estudo foi avaliar o efeito da suplementação de Arg sobre a resposta imune humoral e o desenvolvimento dos órgãos linfoides de frangos de corte imunizados contra a Doença de Gumboro.
Material e métodos
O experimento foi conduzido no Aviário Experimental da Universidade Federal do Paraná – Campus Palotina. Todos os procedimentos de criação dos animais e de coleta de material biológico foram aprovados pelo Comitê de Conduta Ética no Uso de Animais em Experimentação da Federal do Paraná – Campus Palotina, tendo em vista que o ensaio experimental atendeu aos princípios éticos na experimentação animal, conforme preconiza o Colégio Brasileiro de Experimentação Animal.
Foram utilizados 640 pintos de corte, da linhagem Ross, machos, os quais foram alocados aleatoriamente de acordo com um delineamento inteiramente casualizado com quatro tratamentos e oito repetições de 20 aves cada. Os tratamentos foram compostos por: 1. Controle; 2. Imunização contra a Doença de Gumboro; 3. Imunização contra a Doença de Gumboro com suplementação de L-Arg e 4. Suplementação de L-Arg. Para obtenção dos tratamentos com suplementação de L-Arg, foi adicionado 1% de L-Arg à ração em substituição ao inerte (Caulim). As aves dos tratamentos 3 e 4 foram imunizados no 16º dia de idade, contra a Doença de Gumboro, cepa intermediária plus (Shering Plough Animal Health®).
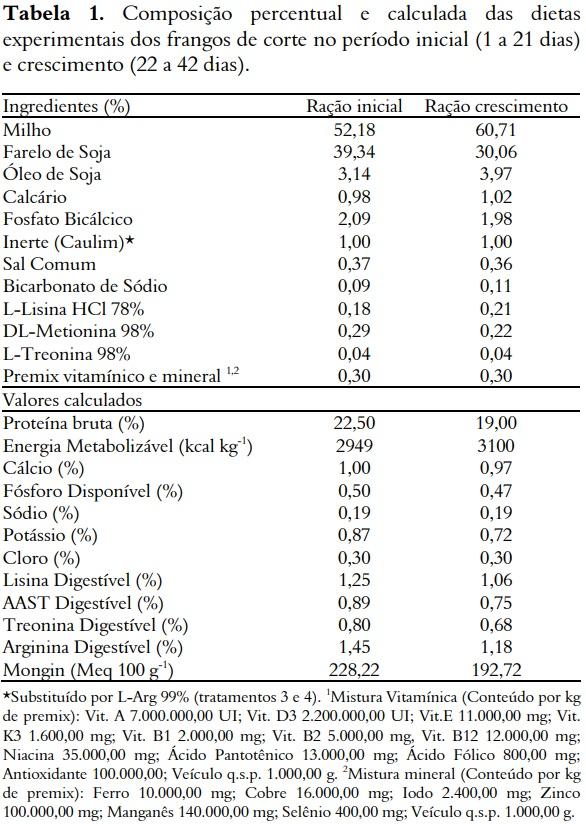
O programa nutricional foi dividido em duas fases, uma inicial (1 a 21 dias) e outra de crescimento (22 a 42 dias). As dietas foram formuladas à base de milho e farelo de soja (Tabela 1), de acordo com os valores de composição química dos alimentos e as recomendações nutricionais propostas por Rostagno et al. (2005).
Arginina e resposta imunológica em frangos
Aos 14, 21, 28 e 35 dias de vida, todas as aves de cada unidade experimental e as sobras de ração foram pesadas, para a determinação do ganho de peso corporal, consumo de alimento e conversão alimentar.
No 14º dia de vida, antes da vacinação, e no 21º, 28º e no 35º foram coletadas amostras de sangue de duas aves por repetição e o soro congelado. Nas amostras de soro foi avaliada a titulação de anticorpos vacinais contra o vírus da Doença de Gumboro, pelo de teste imunoenzimático (ELISA), utilizando o kit comercial ELISA (Enzyme Linked Immuno Sorbent Assay) do Laboratório Idexx® e os dados obtidos foram de absorbância.
Também aos 14, 21, 28 e 35 dias de vida, uma ave por unidade experimental foi sacrificada por deslocamento cervical. Posteriormente, as aves foram pesadas individualmente e removidos os órgãos linfoides (bolsa cloacal e timo). O peso dos órgãos linfoides foi obtido, imediatamente, após a coleta e a remoção dos tecidos exógenos. O peso relativo de cada órgão foi obtido pela seguinte fórmula:
Peso relativo = (peso órgão/peso vivo) x 100
Para a medida histomorfométrica, a bolsa cloacal foi fixada em solução de Bouin e posteriormente inclusa em parafina. Após a obtenção de cortes no centro do órgão com cinco micrômetros de espessura, as lâminas contendo os cortes histológicos foram coradas pelo método da hematoxilina e eosina (HE), escaneadas e levadas ao analisador de imagens. Para a leitura das imagens, foi utilizado um analisador de imagem computadorizado IMAGE PROPLUS 5.2, da Midia Cibertecnics. Foi mensurada a altura de três pregas primárias do tecido linfoide da bolsa cloacal de cada repetição (8 aves por tratamento) e destes valores foi obtida a média.
Os dados de titulação de anticorpos avaliados por ELISA foram transformados em log10. A avaliação estatística dos dados foi feita por análise de variância e comparação das médias pelo teste de Tukey a 5% de probabilidade, utilizando-se o programa SAEG versão 8.0 (SAEG, 1999).
Resultados e discussão
Não houve diferença significativa (p > 0,05) entre os tratamentos para nenhuma variável avaliada (Tabela 2). Provavelmente, o fato das aves vacinadas com vacina intermediária e não-vacinadas deste experimento apresentarem títulos de anticorpos semelhantes, após a vacinação aos 16 dias, se deve à presença de cepas variantes no ambiente onde as aves foram criadas. As cepas variantes têm sido identificadas (BERG, 2000) e é importante destacar que em alguns lotes, a maioria das cepas encontradas nas amostras recebidas para o diagnóstico corresponde a este tipo de cepa. As cepas do vírus que atuam sobre o sistema imunológico dos frangos variam em sua patogenicidade, se encontrando desde cepas vacinais clássicas, que não causam doença, até cepas muito virulentas, que têm a capacidade de produzir porcentagens variáveis de mortalidade, dependendo da imunidade e da idade das aves, quando se atingem as condições do meio ambiente. Também estão presentes as cepas variantes do vírus de Gumboro, que geralmente não causam mortalidade, mas que induzem atrofia severa da bolsa cloacal, que se traduz em menor quantidade de linfócitos e, portanto, menor reação do sistema imunológico da ave (GIAMBRONE; CLOSSER, 1990).
Foi observado neste experimento que houve persistência acima do esperado dos anticorpos maternos. A sorologia antes da vacinação foi feita aos 14 dias e observou-se, nesta data, que o título de anticorpos maternos se mantinha bastante elevado e, por isso, podem ter sido neutralizados pela aplicação da vacina na progênie. Assim, como o agente vacinal, ou de campo, não desenvolveu sintomas ou lesões, com a consequente perda da saúde e rendimento das aves, o nível de Arg dietético necessário para o desempenho das aves foi suficiente para atendimento do sistema imune em fase de ativação.
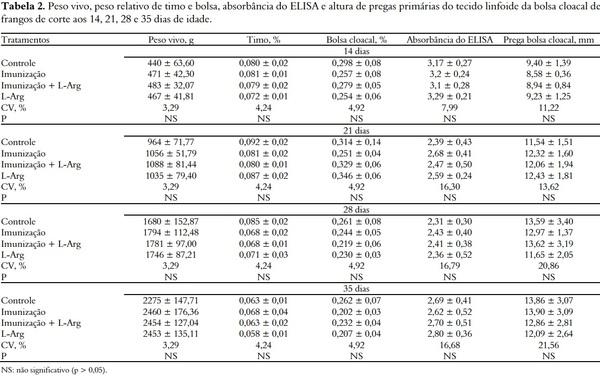
Independente da falta de resposta à vacinação, não foi observado efeito da suplementação de L-Arg, apesar de a mesma estar envolvida em diferentes componentes do sistema imunológico. Este resultado difere do observado por outros autores. Tayade et al. (2006) demonstraram que a resposta vacinal contra o vírus da Doença de Gumboro de frangos suplementados com 2% de Arg foi 20% mais eficiente que a resposta induzida apenas pela vacina sem adição de Arg. A elevada resposta humoral e a proteção observada no grupo de aves vacinadas e que recebeu a suplementação de Arg na dieta, segundo estes autores, pode ser atribuída ao número de funções imunorregulatórias da Arg sobre o sistema imunológico.
A Arg é necessária para a diferenciação das células B e também está envolvida na liberação destas células da bolsa cloacal ou da medula óssea (DE JONGE et al., 2002). Ainda, segundo Abdukalykova e Ruiz-Feria (2006), níveis adicionais de Arg podem acelerar a produção de anticorpos em frangos de corte. Por outro lado, a resposta imune humoral das aves pode ser afetada pela suplementação de Arg apenas em longo prazo. Deng et al. (2005) observaram aumento no título de anticorpos contra hemácias de carneiro, somente após quatro semanas do fim da suplementação de Arg na dieta de frangas de postura.
Outro aspecto a ser considerado, no metabolismo da Arg, é a competição da arginase e a óxido nítrico sintetase pela Arg como um substrato comum, mas que é preferencialmente degradada pela via da arginase I, o que pode levar à redução na síntese de NO pelos leucócitos (WU; MORRIS JR., 1998). Neste sentido, a suplementação adicional de Arg nas dietas de aves imunocomprometidas poderia aumentar ainda mais a atividade da arginase, e, com isso, menos Arg seria disponível para a síntese de NO (KEPKA-LENHART et al. 2000). Alternativamente, Bansal et al. (2004) sugerem como estratégia dietética para restaurar as concentrações plasmáticas de Arg, a suplementação de citrulina, principalmente em condições de imunodeficiência associadas com elevada atividade plasmática da arginase. A citrulina é um aminoácido que não é encontrado em nenhuma proteína corporal, além da inabilidade das aves na síntese endógena de citrulina, a suplementação dietética se constitui na única fonte de Arg para os macrófagos, via ciclo NOcitrulina (WIESINGER, 2001).
Corzo e Kidd (2003) atribuem a exigência mais elevada em Arg, na fase inicial de vida dos frangos de corte, ao desenvolvimento do sistema imune. A Arg estimula a liberação de hormônios, como o hormônio do crescimento, insulina e glucagon, os quais podem aumentar a síntese proteica e a eficiência alimentar. Outro mecanismo inclui à formação de ornitina um precursor das poliaminas, as quais estão envolvidas na síntese de DNA e na proliferação celular e ainda no aumento da formação de NO e na alteração da liberação de citocinas (DENG et al., 2005; KAWAK et al., 1999; LE FLOC’H et al., 2004).
A suplementação com Arg reduziu os efeitos imunossupressores da vacina em outros experimentos. A julgar por estes resultados apontados na literatura consultada, a Arg tem potencial para ser considerada uma substância imunomoduladora segura, pouco dispendiosa e de fácil administração. Entretanto, mais estudos devem ser desenvolvidos para que, efetivamente, possa ser incluída aos programas de vacinação.
Conclusão
A falta de resposta à imunização pode ser atribuída à baixa virulência da cepa vacinal utilizada e à presença de vírus de campo que interferiu na avaliação da resposta imune das aves. A suplementação de Arg não contribuiu com a resposta imune dos frangos de corte vacinados contra a Doença de Gumboro.









.jpg&w=3840&q=75)