Peso Leitão Nascimento
Fatores que influenciam o peso do leitão ao nascimento
Publicado: 14 de agosto de 2012
Por: Andrea Panzardi, Brenda Maria Ferreira Passos Prado Marques, Giseli Heim, Fernando Pandolfo Bortolozzo e Ivo Wentz, Setor de Suínos, Faculdade de Veterinária, Universidade Federal do Rio Grande do Sul (UFRGS), RS.
INTRODUÇÃO.
O peso do leitão ao nascimento (PN) é considerado um dos principais fatores diretamente relacionados à sua sobrevivência [67], bem como com seu peso ao desmame e desempenho posterior, até o momento do abate.
Apesar do grande progresso genético e no manejo ao longo dos anos, a mortalidade pré-desmame de leitões ainda representa um grande entrave econômico na suinocultura moderna [45]. Isto se torna uma realidade quando deparamos com taxas de mortalidade pré-desmame que permanecem relativamente constantes desde meados das décadas de 50 [39], 60 [16], 90 [43] e até hoje em dia [67], girando em torno de 25%, 15%, 20%, 16%, respectivamente, na grande maioria dos plantéis.
Atualmente, as linhagens de fêmeas suínas destinadas à reprodução vêm sendo melhoradas geneticamente, no intuito de tornarem-se hiperprolíficas. Além disso, mudanças inerentes ao manejo possibilitaram o aumento do número de leitões desmamados/fêmea/ano, passando de uma média de 21 a 23 leitões [55] para um patamar em torno de 25 a 28 leitões [65], entretanto, algumas genéticas já atingiram um total de 30 leitões desmamados/ fêmea/ano [4]. Este aumento possibilitou uma melhor produtividade e, consequentemente, maior ganho econômico, uma vez que o valor de cada leitão nascido gira em torno de R$ 30,00 [43]. Como consequência, houve o surgimento de problemas relacionados com o PN e leitegadas desuniformes, contribuindo com uma maior variabilidade de peso entre os leitões. Outro fator importante e intrínseco a este, está relacionado à viabilidade e vitalidade de leitões menos favorecidos, em virtude de seu baixo PN e possível exposição a eventos estressantes durante o parto [17].
Em virtude dos programas de melhoramento genético das empresas serem, primeiramente, focados em características produtivas e reprodutivas, como tamanho da leitegada e ganho de peso diário (GPD) [49], características como capacidade uterina, eficiência placentária e nutrição acabam não sendo focadas na mesma proporção, porém, são de extrema importância para uma perfeita harmonia do processo do desenvolvimento embrionário e fetal como um todo, levando a um bom peso do leitão ao nascer.
IMPORTÂNCIA DO PESO AO NASCER NA SOBREVIVÊNCIA NENONATAL E DESENVOLVIMENTO ATÉ O ABATE
Com os recentes avanços genéticos, hoje é possível atingir um patamar de GPD de leitões em torno de 0,7 kg do desmame ao abate. Dentro das fases de desenvolvimento, o período do nascimento até 30 kg representa a fase na qual o potencial genético máximo do animal ainda não foi alcançado. Neste contexto, um maior desenvolvimento obtido neste período, é em grande parte responsável pelo retorno econômico e pelo sucesso da atividade [20].
Um menor PN predispõe a uma menor chance de sobrevivência [67,82], sendo este efeito verificado em leitões com média de PN inferior a 1,0 kg [67]. Além disso, leitões com baixo PN possuem menores níveis de reservas energéticas corporais, maior sensibilidade ao frio, demoram mais tempo para atingir o complexo mamário e mamar efetivamente, além de terem menor habilidade em escolher os melhores tetos [37,43]. Todos esses fatores em conjunto levam a uma menor ingestão de colostro e leite, menor aquisição de imunidade passiva, gerando um quadro de subnutrição, o que resulta em maior mortalidade pós-natal e comprometimento do desenvolvimento [44,67]. Neste aspecto, fêmeas Meishan aparecem como uma exceção, pois apresentam uma menor taxa de mortalidade de sua progênie, apesar de apresentarem leitões com média de PN inferior aos de fêmeas de outras genéticas (Ex.: 1,32 ± 0,05 vs 1,51 ± 0,04 vs 1,54 ± 0,08, Meishan vs Large White vs Duroc, respectivamente) [11]. Esta melhor viabilidade pode ser devido a uma maior maturidade ao nascimento e, também, possivelmente, uma melhor habilidade no momento do parto, com maior capacidade em expulsar os leitões [37].
Outro fator interessante a ser destacado e associado ao baixo PN é a perda de peso nas primeiras 24 horas de vida, a qual também pode contribuir com a taxa de mortalidade durante o período neonatal [6] (Tabela 1).
Tabela 1. Indicadores de sobrevivência pós-natal (médias e erros padrão) comparando leitões sobreviventes (n=113) e os que morreram (n=13) no período neonatal.
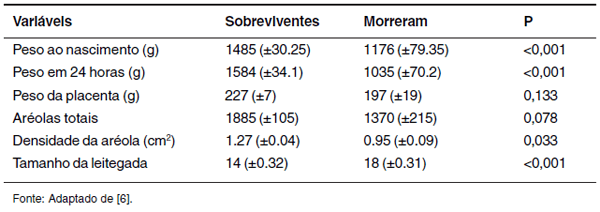
Além disso, a capacidade termorregulatória dos leitões possui grande impacto em sua viabilidade, sendo este parâmetro fisiológico diretamente relacionado ao PN. Leitões mais leves possuem maior superfície corporal em relação ao seu peso, sendo, portanto, mais propensos a um quadro de hipotermia [38]. Panzardi et al. [61] demonstraram que o PN (<1,275kg), a temperatura corporal às 24h pós-nascimento (<38ºC) e a ordem de nascimento (>9) são os melhores indicadores da taxa de sobrevivência durante a primeira semana pós-natal.
Segundo Rehfeldt & Kuhn [71], leitões com baixo, médio e elevado PN apresentaram respectivamente 582, 619, e 641 g de GPD. Este menor GPD em leitões mais leves ao nascimento resulta em um menor peso ao desmame. Isto também foi confirmado por Mahan et al. [51], pois, ao acompanharem leitões desde nascimento até o período de abate, verificaram que aqueles com baixo peso ao desmame (5,5 vs 7,5kg) levaram em média 8 dias a mais para atingir o peso de abate quando comparados a leitões mais pesados (164,8 vs 156,7dias) (P<0,05). Além disso, leitões com baixo PN apresentam um pior rendimento de carcaça, percentual de carne e área muscular de lombo e maior percentual de gordura interna, em relação aos de elevado PN (P=0,01) [71].
1 Peso ao nascer vs maturidade anatomofisiológica
O grau de atraso no desenvolvimento e maturação fetal é um importante fator predisponente para o aumento da mortalidade pré-desmame. Este atraso envolve características como eficiência placentária, maturação funcional dos órgãos vitais e disponibilidade de reservas energéticas [45,72]. Ao comparar leitegadas com mesmo PN, foi observado que aquelas com maior chance de sobrevivência apresentaram peso do fígado, glândulas adrenais, intestino delgado, e estômago (7,1%, 21,7%, 5,4%, e 4,3%, respectivamente) superior às com menor chance, diferindo, apenas, o baço, que apresentou peso 6,1% inferior [45].
O cortisol, por sua vez, estimula e regula a maturação e o desenvolvimento de vários órgãos importantes na sobrevivência após o nascimento [46], além de estimular a deposição de glicogênio muscular e hepático [69] e maturação dos pulmões, trato gastrointestinal e tireóide [46,73]. Assim, as concentrações médias de cortisol são maiores em leitões que apresentam uma maior taxa de sobrevivência [45].
A reserva total de glicogênio corporal em leitões recém-nascidos gira em torno de 30 a 38 g/kg de peso corpóreo [64], a qual se encontra presente no fígado, músculo esquelético e tecido adiposo. Esta reserva cai bruscamente algumas horas após o nascimento, sendo mais acentuada no fígado (70% no primeiro dia de vida). A musculatura esquelética se torna então a principal fonte de energia em longo prazo para a manutenção da homeotermia [38,64]. Leenhouwers et al. [45] verificaram que a concentração de glicogênio hepático e muscular foi, respectivamente, maior (8,1% e 6,2%), em leitões com alta taxa de sobrevivência, sugerindo que estes leitões apresentam uma maior capacidade de manter a glicemia e homeotermia.
FATORES QUE INFLUENCIAM O PESO DO LEITÃO AO NASCER
1 Fatores genéticos
1.1 Hiperprolificidade
Em virtude do intenso processo de melhoramento genético, houve um aumento de produção de uma média de 10,9 leitões nascidos totais na década de 90, para 13,7 em 2006 [76]. Entretanto, com a seleção de fêmeas voltadas para a produção de maior quantidade de leitões por leitegada, houve, consequentemente, a ocorrência da diminuição do PN e um aumento na variação do peso destes [18,56,67,87], como comentado anteriormente. Isto ocorre, pois há uma diminuição do espaço uterino para todos os fetos que se encontram em desenvolvimento. Este evento por sua vez, é determinado ao redor do 15º ao 16º dia de gestação, momento em que há um aumento da associação entre microvilosidades do endométrio e do concepto, através da interdigitação de ambos [19]. Esta adesão entre concepto e endométrio é fundamental para que seja delimitado o espaço uterino disponível para cada concepto competir pelos nutrientes necessários para sua sobrevivência, e assim permitindo um bom desenvolvimento [31].
Esta diminuição do PN foi comprovada por Quiniou et al. [67], que observaram uma redução da média do PN de 1.590 g para 1.260 g quando o tamanho da leitegada aumentou respectivamente de £ 11 para ³ 16 leitões por leitegada, o que representa em média 35 g a menos por leitão a mais nascido.
Amaral Filha [2] verificou que leitoas inseminadas que tinham um maior GPD (701 a 770 g, e 771 a 870 g), quando comparadas àquelas com baixo GPD (600 a 700 g) apresentaram maior tamanho da leitegada, diminuição do PN e, consequentemente, maior coeficiente de variação do PN dos leitões. Isto pode ser explicado pela maior qualidade dos oócitos, permitindo uma maior sobrevivência embrionária.
Um experimento realizado por Roehe [72] demonstrou que a seleção genética para aumento no PN é recomendada, possibilitando uma melhor viabilidade e crescimento pré e pós-natal [87]. Além disso, fatores genéticos foram relacionados à variação de PN dentro de uma mesma leitegada [17,42,72], sendo que Knol et al. [42] observaram que a influência dos genes maternos é muito maior quando comparado aos genes de seus leitões, sendo a herdabilidade para essa mesma característica de 0,005 para efeitos diretos nos leitões, e 0,20 para efeitos genéticos maternos.
Outro fator importante a ser destacado é que, atualmente, a menor deposição de gordura corporal das fêmeas, resulta em menor reserva de gordura durante a fase de gestação, o que poderá culminar em um maior desgaste destas matrizes durante a lactação. Este estado de catabolismo, consequentemente, poderá afetar seu desempenho reprodutivo subsequente [77], podendo comprometer o bom desenvolvimento fetal em sua próxima gestação [8,66].
1.2 Capacidade uterina e eficiência placentária
De acordo com Père et al. [63] e Van Der Lende & Schoenmaker [81], a capacidade uterina pode afetar o tamanho da leitegada e o PN dos leitões. O PN é altamente dependente da quantidade de nutrientes fornecidos através da placenta, sendo influenciado pelo tamanho (massa, superfície) e fluxo sanguíneo [42], características intimamente relacionadas à eficiência placentária (EP).
A EP, por sua vez, é medida através da divisão do PN do leitão pelo peso de sua placenta [86]. Uma EP elevada permitiria que placentas menores fossem capazes de manter o desenvolvimento fetal adequado, sem afetar sua viabilidade [10,85].
A formação da placenta apresenta um desenvolvimento expressivo do 12º a 30º dia de gestação, sendo que após este período a placenta encontra-se completamente formada [24], entretanto, seu crescimento continua até em torno do 70º dia de gestação. Já as membranas embrionárias crescem rápido dentro de um período que vai do 20º a 60º dia de gestação. Posteriormente, esta taxa de crescimento diminui consideravelmente, sendo que ao 70º dia de gestação a placenta e o embrião encontram-se com pesos semelhantes. Neste momento, a placenta cessa seu crescimento, porém os fetos continuam a se desenvolver até o momento do nascimento [53].
Em virtude dos fetos necessitarem de um determinado espaço para seu desenvolvimento e sobrevivência, o comprimento do útero parece ter um papel importante na definição do tamanho da leitegada [9]. No período final do processo de alongamento dos embriões (ao redor de 16 dias) seu comprimento gira em torno de 1 m, entretanto encontram-se enrolados ocupando um espaço de 10 a 20 cm. O alongamento dos embriões cessa no momento em que a placenta de um dos leitões entra em contato com placentas adjacentes, sendo que depois de sete semanas de gestação os fetos necessitam de mais de 20 cm de espaço para sobreviver [88].
Na medida em que o número de embriões não é um fator limitante, uma placenta pouco eficiente passa a ser a principal responsável pelo aumento da taxa de mortalidade pré-natal [85]. Isto se torna uma característica importante, uma vez que o desempenho pós-natal é determinado, em grande parte, pelo desenvolvimento intrauterino [28].
Um estudo associando a variação do PN de leitões, sua sobrevivência pré-desmame e seu GPD, concluiu que a seleção genética para o aumento do tamanho da leitegada, o qual induzirá diminuição do PN e aumento da variação do peso da leitegada não seria o mais indicado. A capacidade uterina se tornaria um fator limitante, a não ser que algumas medidas fossem tomadas no intuito de melhorar a sobrevivência da progênie de baixo PN [56].
A herdabilidade encontrada para a EP é superior à observada para a capacidade uterina ou nascidos totais [79]. Neste sentido, Vonnahme et al. [83] sugerem que a seleção com base nessas características poderia resultar em uma diminuição de perdas fetais, uma vez que fêmeas selecionadas para uma alta EP tiveram 3,3 leitões/ leitegada a mais que as selecionadas para uma baixa EP (12,8 vs. 9,5 leitões) [84].
Ao avaliarem a EP no intuito de verificar o quão melhor esta característica é em relação ao PN e peso de placenta para predizer o risco de mortalidade pré-desmame, Van Rens et al. [82] observaram que a EP é uma característica complicada de ser avaliada, uma vez que o efeito da EP sobre risco de mortalidade pré-desmame é altamente dependente do PN e peso de placenta, sendo que dessas duas características, o PN é visto como o melhor preditor para o efeito da EP sobre a mortalidade pré-desmame.
A estimativa genética de valores para sobrevivência dos leitões está relacionada positivamente com a EP e inversamente com o peso da placenta [45]. Entretanto, há um fator limitante que é determinado pela capacidade uterina, uma vez que a EP aumenta de acordo com o aumento do PN, porém somente até atingir um peso máximo em torno de 1,6 kg declinando, posteriormente [82].
Além da EP, a densidade e o número total de aréolas foram as características placentárias mais importantes no que diz respeito à sobrevivência pós-natal [6] (Tabela 1).Estas estruturas são os principais sítios de transferência de nutrientes no útero, surgem em torno do 30º dia de gestação e apresentam um tamanho individualmente semelhante a uma glândula uterina numa fase mais precoce da gestação. Em uma fase mais avançada, aumentam em tamanho e número, levando a fusão de algumas, formando, então, aberturas glandulares múltiplas [24]. Aréolas mais densas melhoram o ambiente intrauterino, chances de crescimento, desenvolvimento e sobrevivência dos leitões [6].
1.2.1 Diferenças entre raças
Sabe-se da existência de diferenças inerentes às raças no que diz respeito à prolificidade. A raça Meishan, por exemplo, apresenta menor tamanho de placenta e melhor EP quando comparada a raças européias e americanas [9]. Isto significa que os fetos Meishan, por ocuparem um menor espaço uterino, possibilitam uma maior sobrevivência embrionária [25], consequentemente, produzindo um maior número de leitões e mais homogêneos [85].
A EP é maior na raça Meishan, portanto, mesmo que a placenta não cresça muito no final da gestação, ela apresenta uma proliferação significativa de vasos sangüíneos na membrana corio-alantóide, em contraste com a placenta de fetos Yorkshire [10,25]. Ao analisarem o peso da placenta de leitões Meishan e Yorkshire aos 90 dias de gestação e ao parto, Wilson et al. [85] verificaram que placentas de fetos Meishan apresentaram pesos similares tanto aos 90 dias quanto ao parto, já em contrapartida, placentas de fêmeas Yorkshire, apresentaram um aumento de peso em torno de 70% dos 90 dias de gestação até o parto. Fica evidente, então, que nas fêmeas Meishan o fluxo sanguíneo placentário assume grande importância para que a placenta seja mais eficiente na disponibilização de nutrientes para o feto. A adequada angiogênese da placenta é crítica para o estabelecimento da circulação placentária e manutenção de fluxo sanguíneo uterino e umbilical adequados para o crescimento normal dos fetos. Fatores que influenciam os aspectos de desenvolvimento e função da vascularização da placenta podem ter efeito marcante no crescimento fetal e, portanto, com consequências na sobrevivência e crescimento neonatal.
Ao comparar o desempenho de quatro raças (Large White (LW), Meishan (MS), linha macho Laconie (LA) e F1 Duroc X Large White (DU x LW)), foi observado que as fêmeas MS tiveram maior (P<0,01) tamanho de leitegada (13,3 vs 12,2 vs 12,8 vs 11,3 ), menor (P<0,01) número de natimortos (0,3 vs 0,7 vs 0,6 vs 0,7,) e menor (P<0,001) PN (1,32 vs 1,51 vs 1,54 vs 1,53 ) que fêmeas LW, DU x LW e LA, respectivamente [11].
O interesse pela raça Meishan tem sido justificado por sua alta prolificidade, baixo percentual de natimortos e alta taxa de sobrevivência do nascimento ao desmame [33], sendo que atualmente uma das estratégias mais utilizadas para aproveitar a vantagem do alto desempenho reprodutivo dessas fêmeas, tem sido o desenvolvimento de novas genéticas, através do cruzamento entre raças chinesas, européias e americanas [92].
Dois são os aspectos que explicam a maior prolificidade da raça Meishan: o primeiro está relacionado à menor taxa de crescimento inicial, resultando em uma menor produção de estradiol e mudanças menos marcantes e mais graduais na composição do histiotrofo uterino, indicando, então, um maior número de conceptos após os 18 dias de gestação [25,54]; o segundo fator é atribuído a uma placenta menor, de alta EP, ocupando um menor espaço uterino, e não diminuindo a viabilidade fetal [85].
1.3 Restrição do crescimento intra-uterino (IUGR)
A capacidade funcional da placenta é essencial para uma perfeita harmonia do desenvolvimento dos leitões que compõe uma leitegada. Logo após a implantação do embrião, há a expressão de diversos genes, os quais têm como principal função iniciar o processo de placentação [3]. Este processo, por sua vez, envolve uma relação íntima entre placenta e feto, e alguns processos regulatórios devem já estar pré-programados pela ativação destes genes [91].
O desenvolvimento da placenta dos leitões é estimulado por fatores regulatórios do crescimento representados pelo fator de crescimento semelhante à insulina (IGF) e fator de crescimento vascular endotelial (VEGF) [19]. Durante seu desenvolvimento, a placenta forma estruturas que se ligam ao endométrio, formando uma superfície de contato essencial para a troca de nutrientes e gases entre mãe e feto. Além disso, esta superfície promove o desenvolvimento de um fluxo sanguíneo através da vasodilatação de veias placentárias [76]. Uma boa vascularização da placenta é fundamental para que haja uma boa suplementação de aminoácidos aos fetos. Um fator extremamente crítico é a formação de novos vasos sanguíneos a partir de vasos já pré-existentes, denominado angiogênese, que ocorre na parede do endométrio, em resposta a um aumento da demanda de substratos para os tecidos. Em contraste, a vasculogênese ocorre somente nos conceptos e envolve a diferenciação das células mesenquimais em angioblastos [74].
Sabe-se que aminoácidos da família da arginina (arginina, prolina e glutamina) são substratos essenciais para um bom desenvolvimento da placenta e de fetos suínos [89], sendo a arginina precursora do óxido nítrico (NO) e responsável pela síntese de poliaminas (Figura 1). O NO tem um papel extremamente importante, no que diz respeito à vascularização da placenta e na regulação de seu fluxo sanguíneo, sendo, consequentemente, responsável pela transferência de nutrientes e oxigênio (O2) da mãe para o feto. Uma redução em sua síntese e/ou liberação, certamente, levaria a uma alteração deste suprimento. As poliaminas, que são sintetizadas na placenta suína a partir de substratos derivados da prolina, regulam o DNA e síntese protéica, estando diretamente relacionadas à proliferação e diferenciação celular [90]. O NO e as poliaminas são as chaves reguladoras da angiogênese, embriogênese, assim como do crescimento placentário e fetal [89].
Uma vascularização insuficiente da placenta e um menor fluxo sanguíneo leva a uma deteriorização progressiva da função placentária, levando a diminuição da transferência transplacentária de O2 e nutrientes para os fetos, resultando em uma IUGR [30]. Esta IUGR, de modo geral, pode ser definida como um comprometimento no crescimento e desenvolvimento de embriões e/ou fetos de mamíferos durante a gestação [91]. Na espécie suína, trata-se de um fenômeno comum em fêmeas hiperprolíficas, uma vez que há uma intensa disputa intrauterina entre os fetos. Fêmeas Meishan aparecem como uma exceção no que diz respeito à ocorrência da IUGR, pois apesar de serem hiperprolíficas e apresentarem, relativamente, uma placenta mais leve, elas apresentam um alto grau de vascularização da mesma, o que permite uma melhor troca materno-fetal [25].
Figura 1. Mecanismo pelo qual a arginina promove o crescimento placentário e fetal. Fonte: Adaptado de [89].
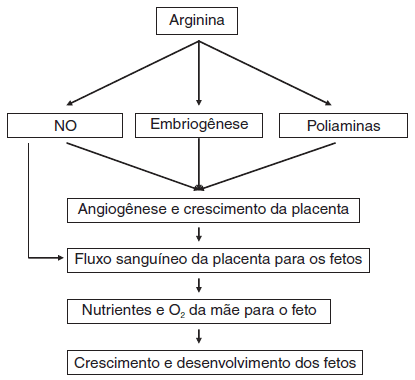
O mecanismo que leva a essa insuficiência placentária e, consequentemente, à IUGR foi revisado por Wu et al. [91], e envolve uma interação complexa entre genética, epigenética, fatores ambientais, bem como a maturidade materna. Estes mesmos autores sugerem que a trajetória do crescimento da placenta e do feto é naturalmente vulnerável a uma subnutrição, supernutrição, estresse térmico, doenças e toxinas durante a gestação, porém, a IUGR é potencializada caso este distúrbio nutricional ocorra no período em que há um máximo crescimento da placenta.
Muitas leitegadas apresentam ao menos um leitão com baixo PN, sendo que na maioria dos casos, este baixo PN se trata de um exemplo da ocorrência da IUGR [5]. Essa IUGR gerada, como já dito anteriormente, resulta em menor PN [20,51,71], o qual está diretamente relacionado à sua sobrevivência, peso ao desmame e desempenho subsequente [67]. Este menor PN contribui com 37% na variação do peso ao desmame [15]. Dados de Furtado [29] confirmam tais resultados, uma vez que houve uma correlação positiva (r= 0,515; P<0,001) entre o PN e ao desmame, e, além disso, verificaram que para cada grama a mais ao nascimento houve um incremento de 2 g no peso ao desmame.
1.4 Miogênese
Nas espécies multíparas, como na suína, há naturalmente uma variação no PN, sendo esta variação fortemente relacionada com o número de fibras musculares presentes. Leitões com menor PN apresentam um menor número de fibras musculares, sendo decorrente de um menor número de fibras que se diferenciaram durante o período de miogênese pré-natal, por motivos genéticos (cruzamento, genótipo) ou maternos (nutrição). Este número de fibras reduzido torna estes leitões menos capazes de apresentar uma recuperação em termos de desempenho e ganho de peso no período pós-nascimento [71].
1.4.1 Formação das fibras musculares dos leitões durante a gestação e desempenho pós-nascimento
O desenvolvimento das fibras musculares dos leitões durante a gestação ocorre em duas etapas distintas (Figura 2) [7]:
- Formação primária: Os formadores das primeiras fibras musculares pré-natais são denominadas de fibras musculares primárias [34]. Uma vez formadas, elas atuam como estruturas nas quais os mioblastos se unem para formar as fibras musculares secundárias. Este tipo de fibra é influenciado diretamente pela genética (não afetada por nutrição ou condições do ambiente uterino).
Figura 2. Representação esquemática dos períodos de desenvolvimento das fibras musculares nos fetos suínos. Fonte: Adaptado de [28].
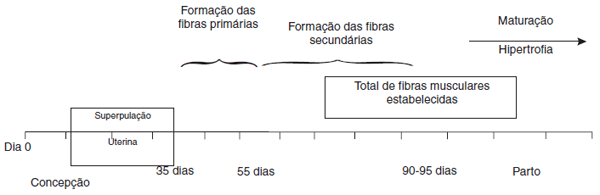
- Formação secundária: Esta segunda formação engloba o período de hiperplasia das fibras musculares secundárias (25 a 90 dias de gestação) [26]. Esta fase é determinada por eventos pré-natais, relacionados, principalmente, a fatores nutricionais e limitação do espaço uterino da fêmea durante o desenvolvimento fetal.
Estudos revelaram a presença de quatro grupos de fatores reguladores da miogênese (MRFs): myoD, myf5, myogenin e MRF4 (myf6), sendo que todos podem ativar a diferenciação da musculatura esquelética. Estes MRFs consistem num grupo de fatores de transcrição responsáveis pela regulação central do programa de desenvolvimento muscular esquelético [52] Experimentos demonstraram que o myoD possui uma função principal na formação e na sobrevivência dos mioblastos, desempenhando um importante papel na diferenciação do mioblasto, enquanto acredita-se que o myogenin está envolvido na diferenciação final em miotubos [70].
O processo de miogênese, por sua vez, pode ser afetado pela superlotação uterina. Diferenças relacionadas a esta superlotação durante o período pré-natal levam a consequências no padrão do desenvolvimento das fibras musculares [26] e, consequentemente, causando uma IURG. Num experimento desenvolvido por Town et al. [78] foi verificado um efeito moderado do processo de superlotação uterina na expressão de fatores reguladores da miogênese, como myogenin e myoD. Os autores concluíram que a superlotação intrauterina aos 30 dias de gestação pode causar um impacto na diferenciação das fibras musculares em virtude da redução da expressão da myogenin.
Estes resultados demonstrados acima sugerem que alterações no ambiente uterino durante períodos precoces da gestação podem comprometer o bom desenvolvimento embrionário e fetal levando a diferenças inerentes ao peso da leitegada e, consequentemente, prejudicando o desempenho pós-natal do leitão.
2 Fatores nutricionais
A nutrição da fêmea suína é uma área bastante explorada e de constantes desafios no sentido de atender matrizes atualmente mais precoces, mais produtivas e, nutricionalmente mais exigentes. As práticas de alimentação das diferentes categorias de fêmeas embora tenham objetivos específicos, estão interrelacionadas, o que faz com que o programa de nutrição em uma determinada fase tenha efeitos significativos no desempenho alcançado na fase subsequente [1]. Todas as fases da gestação são importantes quanto à nutrição, sendo necessário adequar os níveis nutricionais para cada uma delas, pois o alimento é fornecido de forma restrita. O não cumprimento destas exigências influencia a taxa de crescimento e desenvolvimento dos fetos no útero e consequentemente o PN [13]. Na espécie suína, uma suplementação inapropriada de nutrientes no útero, resulta em 15 a 20% de leitões com baixo PN, cuja sobrevivência e desenvolvimento pós-natal já estarão comprometidos [64]. Os efeitos nutricionais e diferentes estratégias utilizadas diferem com relação à fase gestacional em que a fêmea se encontra. Com isso temos, teoricamente, que o período de gestação da fêmea suína pode ser dividido em 3 fases:
2.1 Fase inicial da gestação (primeiros 21 dias)
Esta fase caracteriza-se pela ligação embrio-maternal, início da formação da placenta e anexos fetais, o que exige, portanto, menor necessidade de ganho de peso e reserva energética da fêmea. Neste período, tanto uma subnutrição como uma supernutrição pode ser prejudicial. Uma alimentação deficiente pode resultar em menor síntese de NO e de poliaminas, o que resulta, como já dito anteriormente, em menor vascularização placentária e transferência de nutrientes da mãe ao feto. Isto leva à subnutrição fetal, comprometendo o crescimento intrauterino [89].
Por outro lado, altos níveis de consumo alimentar durante o início da gestação também podem exercer influência negativa sobre a sobrevivência embrionária. Isto é atribuído à redução da concentração de progesterona plasmática, devido ao aumento do fluxo sanguíneo e do catabolismo hepático deste hormônio, causados pelo alto consumo de alimento [18]. Por sua vez, a progesterona influencia as atividades secretórias do útero e do oviduto necessárias para o embrião em desenvolvimento [27]. O período crítico para sobrevivência embrionária compreende as primeiras 48 e 72 horas da gestação, onde então, recomenda-se limitar o consumo de ração [40]. A condição corporal ou o estado energético da fêmea influencia a resposta desses animais a altos níveis de consumo alimentar, sendo que a mortalidade embrionária somente aumenta quando altos níveis de alimentos são fornecidos a fêmeas que apresentam boas condições corporais [1].
2.2 Fase intermediária da gestação (22 a 75 dias)
Trata-se de uma fase da gestação em que há a recuperação das reservas corporais das fêmeas, mobilizadas na lactação anterior. A nutrição após o período crítico inicial da prenhez e até o início do terço final da gestação influencia mais a composição corporal da fêmea do que o tamanho da leitegada ou o peso dos leitões [13], apesar de ser nessa fase em que há o estabelecimento do número de fibras musculares nos fetos.
Em virtude da hiperplasia das fibras musculares já estar cessada no momento do nascimento, o nível de hipertrofia e a massa corpórea final dependerá do número de fibras musculares formadas durante o desenvolvimento fetal. Acredita-se que a heterogeneidade dentro de uma mesma leitegada esteja relacionada a uma baixa taxa de crescimento das fibras musculares secundárias [21,26]. Segundo alguns estudos, uma subnutrição uterina durante este período poderia afetar o PN, pela redução do número de fibras musculares secundárias. No entanto, os trabalhos sobre a influência da alimentação materna vs miogênese dos fetos tem apresentado resultados contraditórios.
Ao avaliarem o crescimento de leitões desde o nascimento até atingirem 80 kg de peso vivo, Dwyer et al. [21] observaram uma correlação positiva do GPD, entre 25 e 80 kg, com o número total de fibras musculares presentes no músculo semitendinoso ao nascimento. Embora a nutrição materna no terço final da gestação possa afetar o PN, o número de fibras musculares pode ser alterado por aumento no nível nutricional materno, em fase mais precoce da gestação.
Um efeito mais pronunciado no número de fibras secundárias foi observado quando o aumento no nível nutricional foi efetuado no período que precede a hiperplasia das fibras secundárias (25 - 50 dias de gestação) [22]. Por outro lado, Musser et al. [57] e Nissen et al. [59], ao aumentarem o consumo energético das fêmeas, entre os dias 29 e 45 e 25 e 50 de gestação, respectivamente, não encontraram algum efeito sobre o número de fibras musculares e peso dos fetos. Mais recentemente, Bee [7] verificou que a alimentação não influenciou o número de fibras musculares, embora tenha alterado o tipo de fibra muscular.
O efeito da utilização de somatotropina (pST) e L-carnitina tem sido avaliado em vários estudos. A administração de pST em fêmeas entre os dias 28 e 42 dias de gestação reduziu a variação de peso dos fetos [75], enquanto, que leitões de fêmeas que receberam L-carnitina na gestação apresentaram maior taxa de crescimento durante o período de amamentação do que leitões do grupo controle [23,68]. Fêmeas alimentadas com L-carnitina durante a gestação (5º ao 112º dia) apresentaram maiores concentrações de insulina e IGF-I entre 60 e 90 dias, período de desenvolvimento das fibras musculares secundárias do feto. A adição de L-carnitina na dieta melhorou a utilização dos nutrientes dietéticos, resultando em um aumento do peso das reservas de gordura e um maior PN. Os efeitos positivos da administração de pST e L-carnitina [58] estariam relacionados a possível elevação dos níveis de IGF-I, o que, segundo Magri et al.[50], seria importante na proliferação e diferenciação das células miogênicas, melhorando o crescimento pós-natal dos leitões.
2.3 - Fase final da gestação (76 dias até o parto)
Nesta fase a necessidade de ganho e reserva energética torna-se, expressivamente, maior quando comparada aos dois períodos anteriores, uma vez que representa a fase de maior intensidade de crescimento fetal. O aumento do consumo energético e protéico no último terço de gestação pode aumentar o PN. Este efeito pode ser de vital importância para as linhagens hiperprolíficas, com tendência a um maior crescimento materno e maior número de leitões de baixo PN. Entretanto, o excesso de energia, entre os 75 e 90 dias de gestação pode resultar em prejuízo na formação da glândula mamária e, consequentemente, menor produção de leite durante a lactação, especialmente em primíparas [36].
Fêmeas alimentadas com 1,82 kg/dia (5.900 Kcal/dia) em relação a fêmeas que recebiam 2,27 kg/dia (7.400 Kcal/dia) apresentaram leitegadas mais pesadas [14]. Por outro lado, King et al. [41] observaram que o aumento da quantidade de ração fornecida e do teor de proteína da dieta entre os 66 e 101 dias de gestação não influenciaram significativamente o PN. De acordo com Close & Cole [13], há um acréscimo de 0,8 kg em cada leitão para cada aumento de 2.390 Kcal ED/dia ingerida. Entretanto, quando o PN do leitão for acima de 1,5 kg, não há resposta significativa a níveis de ingestão de energia acima de 7.500 Kcal ED/dia [41].
Deve ser considerado que o maior fornecimento de energia durante a gestação pode resultar em um menor consumo de ração durante a lactação. Portanto, se faz necessária uma perfeita integração entre essas duas fases, para que seja alcançado um melhor desempenho reprodutivo das matrizes, e, consequentemente, uma maior longevidade das mesmas dentro do plantel reprodutivo.
Com a intensa seleção genética para deposição de massa muscular, tem-se verificado que o metabolismo protéico em fêmeas gestantes e suas exigências de aminoácidos essenciais alteraram. Lima et al. [48] avaliaram os efeitos da ingestão de diferentes níveis (10,0; 13,5 e 17,0) de PB durante a gestação, do primeiro ao terceiro parto, sobre os desempenhos produtivo e reprodutivo. O aumento do peso das fêmeas a cada ciclo consecutivo influenciou o PN médio da leitegada e o peso ao desmame, independentemente do nível de PB da ração de gestação. Por outro lado, Haley et al. [33] observaram aumento no peso dos leitões ao nascimento ao fornecerem níveis crescentes de PB na ração de gestação.
O perfeito entendimento das quantidades diárias de nutrientes necessários para o máximo desempenho de fêmeas e o controle constante do consumo diário de nutrientes possibilitam a formulação adequada de rações para garantia do sucesso na produção de suínos [47]. No entanto, existem poucos estudos sobre o efeito da nutrição em fêmeas suínas de alto potencial e estes apresentam resultados variáveis, principalmente quando considerados vários ciclos reprodutivos.
CONSIDERAÇÕES FINAIS
O peso do leitão ao nascimento é um fator extremamente importante, inicialmente, para sua sobrevivência, e, posteriormente, para um bom desempenho até o momento de abate.
Fêmeas hiperprolíficas produzem um maior número de leitões nascidos por leitegada, o que resulta em menor peso médio ao nascimento e, consequentemente, maior variabilidade de peso desses leitões. A capacidade uterina entra, neste aspecto, como um fator limitante, uma vez que se trata de uma característica, anteriormente já explorada em processos de melhoramento genético na tentativa de aumentar o tamanho da leitegada.
Portanto, características como eficiência placentária e tamanho de placenta devem ser incluídas em programas de melhoramento genético, uma vez que se tratam de aspectos vistos como essenciais para a produção de leitões mais homogêneos e, consequentemente, de melhor viabilidade. A nutrição e o manejo alimentar por sua vez, devem ser realizados minuciosamente, sempre respeitando as diferentes fases reprodutivas em que as fêmeas se encontram, no intuito de proporcionar uma alta qualidade nutricional e assim um bom peso do leitão ao nascimento. Esses dois fatores associados, genética e nutrição, certamente, auxiliarão no aumento da sobrevivência dos leitões durante a fase de lactação e com isso, contribuindo positivamente com a diminuição da taxa de mortalidade na maternidade e com o aumento do ganho econômico da produção.
REFERÊNCIAS
1 Abreu M.L.T., Donzele J.L. & Oliveira R.F.M. 2005. Exigências e manejo nutricionais de matrizes suínas gestantes e lactantes. In: Anais do IV Seminário Internacional de Aves e Suínos (Florianópolis, Brasil). pp.33-59.
2 Amaral Filha W. 2009. Reflexo da taxa de crescimento e do peso corporal da leitoa sobre o desempenho reprodutivo subsequente e longevidade da matriz. 32f. Porto Alegre, RS. Tese (Doutorado em Ciências Veterinárias) - Programa de Pós-graduação em Ciências Veterinárias, Universidade Federal do Rio Grande do Sul.
3 Angiolini E., Fowden A., Coan P., Sandovici I., Smith P., Dean W., Burton G., Tycko B., Reik W., Sibley C. & Constância M. 2006. Regulation of Placental Efficiency for Nutrient Transport by Imprinted Genes. Placenta. 27 (Suppl A): 98-102.
4 Antunes R.C. 2007. Planejando a reposição de reprodutores (macho e fêmea) e impacto sobre a eficiência reprodutiva da granja. Revista Brasileira de Reprodução Animal. 31: 41-46.
5 Ashworth C.J., Finch A.M., Page K.R., Nwagwu M.O. & McArdle H.J. 2001. Causes and consequences of fetal growth retardation in pigs. Reproduction Supplement. 58: 233-246.
6 Baxter E.M., Jarvis S., D'Eath R.B., Ross D.W., Robson S.K., Farish M., Nevison I.M., Lawrence A.B. & Edwards S.A. 2008. Investigating the behavioural and physiological indicators of neonatal survival in pigs. Theriogenology. 69: 773–783.
7 Bee G. 2004. Effect of early gestation feeding, birth weight, and gender of progeny on muscle fiber characteristics of pigs at slaughter. Journal of Animal Science. 82: 826-836.
8 Beltranena E., Aherne F.X., Foxcroft G.R. & Kirkwood R.N. 1991. Effects of pre and post pubertal feeding on production traits at first and second estrus. Journal of Animal Science. 69: 886-893.
9 Bernardi M.L., Wentz I. & Bortolozzo F.P. 2006. Desenvolvimento do concepto suíno e fatores que predispõem à mumificação. In: Anais do I Simpósio UFRGS sobre Produção, Reprodução e Sanidade Suína (Porto Alegre, Brasil). pp.236-250.
10 Biensen N.J., Wilson M.E. & Ford S.P. 1998. The impact of either a Meishan or Yorkshire uterus on Meishan or Yorkshire fetal and placental development to days 70, 90 and 110 of gestation. Journal of Animal Science. 76: 2169-2176.
11 Canario L., Cantoni E., Le Bihan E., Caritez J.C., Billon Y., Bidanel J.P. & Foulley J.L. 2006. Between-breed variability of stillbirth and its relationship with sow and piglet characteristics. Journal of Animal Science. 84: 3185–3196.
12 Casellas J., Noguera J.L., Varona L., Sánchez A., Arque M. & Piedrahita J. 2004. Viability of Iberian x ?Meishan F2 newborn pigs. II. Survival analysis up to weaning. Journal of Animal Science. 82: 1925–1930.
13 Close W.H. & Cole D.J.A. 2001. Nutrition of sows and boars. 1.ed. Nottinghan: Nottinghan University Press, 377p.
14 Coffey M.T., Diggs B.G., Handlin D.L., Knabe D.A., Maxwell C.V., Noland Júnior P.R., Prince T.J. & Gromwell G.I. 1994. Effects of dietary energy during gestation and lactation on reproductive performance of sows: a coperative study. Journal of Animal Science. 72: 4-9.
15 Cole M. & Varley M. 2000. Weight watchers from birth. Pig international. 30: 13-16.
16 Curtis S.E., Heidenreich C.J. & Martin T.G. 1967. Relationship between bodyweight and chemical composition of pigs at birth. Journal of Animal Science. 26: 749-751.
17 Damgaard L.H., Rydhmer L., Lovendahl P. & Grandinson K. 2003. Genetic parameters for within-litter variation in piglet birth weight and change in within-litter variation during suckling. Journal of Animal Science. 81: 604–610.
18 Den Hartog L.A., Vesseur P.C. & Kemp B.V. 1994. Nutrition-reproduction interactions in sows. In: Cole D.A., Wiseman J. & Valey M.A. (Eds). Principles of pig science. Nottingham: Nottingham University Press, p.215.
19 Dantzer V. & Winther H. 2001. Histological and immunohistochemical events during placentation in pigs. Reproduction. 58 (Suppl): 209- 222.
20 Donzele J.L., Abreu M.L.T. & Hannas M.I. 2002. Recentes avanços na nutrição de leitões. In: Anais do Simpósio sobre Manejo e Nutrição de Aves e Suínos e Tecnologia da Produção de Rações (Campinas, Brasil). pp.103-161.
21 Dwyer C.M., Fletcher J.M. & Stickland N.C. 1993. Muscle Cellularity and Postnatal growth in the pig. Journal of Animal Science. 71: 3339-3343.
22 Dwyer C.M., Stickland N.C. & Fletcher J.M. 1994. The influence of maternal nutrition on muscle fiber number development in the porcine fetus and on subsequent postnatal growth. Journal of Animal Science. 72: 911-917.
23 Eder K., Ramanau A. & Kluge H. 2001. Effect of l-carnitine supplementation on performance parameters in gilts and sows. Journal of Animal Physiology and Animal Nutrition. 85: 73-80.
24 Echeverri H.M. 2004. Selection for placental efficiency in swine. 106f. Columbia, U.S.A. PhD Tesis (Faculty of the Graduate School) - University of Missouri-Columbia.
25 Ford S.P. 1997. Embryonic and fetal development in different genotypes in pigs. Journal of Reproduction and Fertility. 52 (Suppl): 165-176.
26 Foxcroft G.R., Dixon W.T., Novak S., Putman C.T., Town S.C. & Vinsky M.D.A. 2006. Prenatal programming of postnatal growth performance. In: University of Minnesota Reproduction Workshop: Achieving and Exceeding Sow Production Targets (Alberta, Canada). pp.57-72.
27 Foxcroft G.R., Dixon W.T., Treacy B.K., Jiang L., Novak S., Mao J. & Almeida F.C.L. 2000. Insights into conceptus-reproductive tract interactions in the pig. Journal of Animal Science. 77: 1-15.
28 Foxcroft G.R. & Town S. 2004. Prenatal programming of postnatal performance – The unseen cause of variance. Advanced Pork Production. 15: 269–279.
29 Furtado C.S.D. 2007. Influência do peso ao nascimento e lesões no desempenho de leitões lactentes. 45f. Porto Alegre, RS. Dissertação (Mestrado em Ciências Veterinárias) - Programa de Pós-graduação em Ciências Veterinárias, Universidade Federal do Rio Grande do Sul.
30 Gagnon R. 2003. Placental insufficiency and its consequences. Reproductive Biology. 110 (Suppl): 99–107.
31 Geisert R.D. & Yelich J.V. 1997. Regulation of conceptus development and attachment in pigs. Journal of Reproduction and Fertility. 52 (Suppl):133-149.
32 Gluckman P.D. & Harding J.E. 1997. The physiology and pathophysiology of intrauterine growth retardation. Hormone Research. 48 (Suppl): 11–16.
33 Haley C.S., Lee G.L. & Ritchie M. 1995. Comparative farrowing to weaning performance in Meishan and Large White pigs and crosses. Journal of Animal Science. 60: 259–267.
34 Handel S.E. & Stickland N.C. 1987. The growth and differentiation of porcine sckeletal muscle fibers types and the influence of birthweight. Journal of Anatomy Veterinary. 152: 107-119.
35 Hashimoto F.A.M., Ferreira A.S., Donzele J.L., Lima K.R.S., Paiva A.L.C. & Torres C.A.A. 2004. Níveis de Proteína Bruta na Ração de Gestação para Porcas de Segundo e Terceiro Ciclos Reprodutivos. Revista Brasileira de Zootecnia. 33: 365-374.
36 Head R.H. & Williams I.H. 1991. Mammogenesis is influenced by pregnancy nutrition. In: Batterham E.S. (Ed). III Manipulating Pig Production. Qttword: Australasian Pig Science Association, p.76.
37 Herpin P., Le Dividich J., Hulin J.C., Fillaut M., de Marco F. & Berlin R. 1996. Effects of the level of asphyxia during delivery on viability at birth and early postnatal vitality of newborn pigs. Journal of Animal Science. 74: 2067–2075.
38 Herpin P., Damon M. & Le Dividich J. 2002. Development of thermoregulation and neonatal survival in pigs. Livestock Production Science. 78: 25–45.
39 Hutchinson H.D., Terrill S.W., Morrill C.C., Norton H.W., Mead R.J., Jensen A.H. & Becker D.E. 1954. Causes of baby pig mortality. Journal of Animal Science. 13: 1023.
40 Jindal R., Cosgrove J.R., Aherne F.X. & Foxcroft G.R. 1996. Effect of nutrition on embryo mortality in gilts: association with progesterone. Journal of Animal Science. 74: 620.
41 King R.H., Eason P.J., Smits R.J., Morley W.C. & Henman D.J. 2006. The response of sows to increased nutrient intake during mid to late gestation. Australian Journal of Agricultural Research. 57: 33-39.
42 Knol E.F., Leenhouwers J.I. & Van Der Lende T. 2002. Genetic aspects of piglet survival. Livestock Production Science. 78: 47–55.
43 Lay Júnior D.C., Matteri R.L., Carroll J.A., Fangman T.J. & Safranski T.J. 2002. Preweaning survival in swine. Journal of Animal Science. 80: 74–86.
44 Le Dividich J. & Noblet J. 1981. Colostrum Intake and thermoregulation m the neonatal pig relation to environmental temperature. Biology of the Neonate. 40: 167-174.
45 Leenhouwers J.I., Knol E.F., De Groot P.N., Vos H. & Van Der Lende T. 2002. Fetal development in the pig in relation to genetic merit for piglet survival. Journal of Animal Science. 80: 1759-1770.
46 Liggins G.C. 1994. The role of cortisol in preparing the fetus for birth. Reproduction, Fertility and Development. 6: 141–150.
47 Lima G.J.M.M. & Viola E. 1998. Nutrição de porcas em gestação e lactação: qual a sua influência sobre o desenvolvimento da leitegada? In: Anais do Simpósio Sobre Nutrição e Manejo de Leitões (Campinas, Brasil). p.102.
48 Lima K.R.S., Ferreira A.S., Donzele J.L., Manno M.C., Araújo D., Rostagno H.S. & Silva F.C.O. 2006. Desempenho de porcas alimentadas durante a gestação, do primeiro ao terceiro parto, com rações com diferentes níveis de proteína bruta. Revista Brasileira de Zootecnia. 35: 1999-2006.
49 Løvendahl P., Damgaard L.H., Nielsen B.L., Thodberg K., Su G. & Rydhmer L. 2005. Aggressive behaviours of sows at mixing and maternal behaviour are heritable and genetically correlated traits. Livestock Production Science. 93: 73-85.
50 Magri K.A., Ewton D.Z. & Florini J.R. 1991. The role of the IGFs in myogenic differentiation. In: Raizada M.K. & Le Roith D. (Eds). Molecular Biology and Physiology of Insulin-like Growth Factors. New York: Plenum Press, pp.57-76.
51 Mahan D.C., Cromwell G.L., Ewan R.C., Hamilton C.R. & Yen J.T. 1998. Evaluation of the feeding duration of phase1 nursery diet to three-week-old pigs of two weaning weights. Journal of Animal Science. 76: 578-583.
52 Maltin C.A., Delday M.I., Sinclair K.D., Steven J. & Sneddon A.A. 2001. Impact of manipulations of myogenesis in utero on the performance of adult skeletal muscle. Reproduction. 122: 359-374.
53 Marrable A.W. 1971. The embryonic pig. A chronological account. London: Pitman Medical, 130p.
54 Meredith M.J. 1995. Pig breeding and fertility. In: Meredith M.J. (Ed). Animal breeding and infertility. Oxford: Blackwell Science, pp.278-353.
55 Merks J., Ducro-Steverink D. & Feitsma H. 2000. Management and Genetic factors affecting fertility in Sows. Reproduction of Domestical Animal. 35: 261-266.
56 Milligan B.N., Dewey C.E. & De Grau A.F. 2002. Neonatal-piglet weight variation and its relation to pre-weaning mortality and weight gain on commercial farms. Precentive Veterinary Medicine. 56: 119-127.
57 Musser R.E., Davis D.L., Goodband R.D., Tokach M.D. & Nelssen J.L. 1997. Fetal and maternal responses to feed intake from d 29 to 45 of gestation. Journal of Animal Science. 75 (Suppl 1): 165.
58 Musser R.E., Goodband R.D., Tokach M.D., Owen K.Q., Nelssen J.L., Blum S.A., Dritz S.S. & Civis C.A. 1999. Effects of Lcarnithine fed during gestation and lactation on sow and litter performance. Journal of Animal Science. 77: 3289-3295.
59 Nissen P.M., Danielsen V.O., Jorgensen P.F. & Oksbjerg N. 2003. Increased maternal nutrition of sows has no beneficial effects on muscle fiber number or postnatal growth and has no impact on the meat quality of the offspring. Journal of Animal Science. 81: 3018-3027.
60 Noblet J., Dourmad J.Y., Etienne M. & Le Dividich J. 1997. Energy metabolism in pregnant sows and newborn pigs. Journal of Animal Science. 75: 2708-2714.
61 Panzardi A., Bierhals T., Mellagi A.P.G., Bernardi M.L., Bortolozzo F.P. & Wentz I. 2009. Survival of piglets according to physiological parameters at birth. In: Proceedings of the 8th International Conference on Pig Reproduction (Banff, Canada). [in press].
62 Pardi G., Cetin I., Marconi A.M., Lanfranchi A., Bozzetti P., Ferrazzi E., Buscaglia M. & Battaglia F.C. 1993. Diagnostic value of blood sampling in fetuses with growth retardation. The New England Journal of Medicine. 328: 692–696.
63 Père M.C., Dourmad J.Y. & Etienne M. 1997. Effect of number of pig embryos in the uterus on their survival and development and on maternal metabolism. Journal of Animal Science. 75: 1337-1342.
64 Pettigrew J.E. 1981. Supplemental dietary fat for peripartal sows: A Review. Journal of Animal Science. 53: 107-117.
65 PigChamp incorporated company knowledge software. 2008. Resultado da comparação de dados de 2007. Disponível em: <http:/ /www.agrocerespic.com.brhttp://images.engormix.com/arqDownload/278Comparacao%202007.pdf>. Acessado em 08/2008.
66 Prunier A. & Quesnel H. 2000. Nutritional influences on the hormonal control of reproduction in female pigs. Livestock Production Science. 63: 1–16.
67 Quiniou N., Dagorn J. & Gaudré D. 2002. Variation of piglet's birth weight and consequences on subsequent performance. Livestock Production Science. 78: 63–70.
68 Ramanau A., Kluge H., Spike J. & Eder K. 2004. Supplementation of sows with L-carnithine during pregnancy and lactation improves growth of the piglets during the suckling period through increased milk production. Journal Nutritional. 134: 86-92.
69 Randall G.C.B. 1988. Tissue glycogen concentrations in hypophysectomized pig fetuses following infusion with cortisol. Journal of Developmental Physiology. 10: 77–83.
70 Rawls A., Wilson-Rawls J. & Olson N. 1995. Genetic regulation of somite formation. Current Topics in Developmental Biology. 47: 131– 154.
71 Rehfeldt C. & Kuhn G. 2006. Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis. Journal of Animal Science. 84 (Suppl): 113–123.
72 Roehe R. 1999. Genetic determination of individual birth weight and its association with sow productivity traits using Bayesian analyses. Journal of Animal Science. 77: 330-343.
73 Sangild P.T., Fowden A.L. & Trahair J.F. 2000. How does the foetal gastrointestinal tract develop in preparation for enteral nutrition after birth? Livestock Production Science. 66: 141–150.
74 Sherer D.M. & Abulafia O. 2001. Angiogenesis during implantation, and placental and early embryonic development. Placenta. 22: 1-13.
75 Sterle J.A., Cantley T.C., Lamberson W.R., Lucy M.C., Gerrard D.E., Matteri R.L. & Day B.N. 1995. Effects of recombinant porcine somatotropin on placental size, fetal growth, and IGF-I and IGF-II concentrations in pigs. Journal of Animal Science. 73: 2980-2985.
76 Smits C.H.M., Ramaekers P., Kemp B., Hazeleger W. & Wu G. 2006. The role of functional nutrients in prenatal survival and growth of porcine fetuses in early gestation. In: Proceedings of University of Minnesota Reproduction Workshop: Achieving and Exceeding Sow Production Targets (Alberta, Canada). pp.57-72.
77 Thaker M.Y.C. & Bilkei G. 2005. Lactation weight loss influences subsequent reproductive performance of sows. Animal Reproduction Science. 88: 309-318.
78 Town S.C., Putman C.T., Turchinsky N.J., Dixon W.T. & Foxcroft G.R. 2004. Number of conceptuses in utero affects porcine fetal muscle. Reproduction. 128: 443–454.
79 Vallet J.L., Leymaster K.A., Cassady J.P. & Christenson R.K. 2001. Are hematocrit and placental selection tools for uterine capacity in swine? Journal of Animal Science. 79: 64.
80 Van Der Lende T., Knol E.F. & Leenhouwers J.I. 2001. Prenatal development as a predisposing factor for perinatal losses in pigs. Reproduction Supply. 58: 247–61.
81 Van Der Lende T. & Schoenmaker G.J.W. 1990. The relationship between ovulation rate and litter size before and after day 35 of pregnancy in gilts and sows: an analysis of published data. Livestock Production Science. 26: 217-229.
82 Van Rens B.T.T.M., De Koning G., Bergsma R. & Van Der Lende T. 2005. Preweaning piglet mortality in relation to placental efficiency. Journal of Animal Science. 83: 144–151.
83 Vonnahme K.A., Wilson M.E., Foxcroft G.R. & Ford S.P. 2002. Impacts on conceptus survival in a commercial swine herd. Journal of Animal Science. 80: 553-559.
84 Wilson M.E., Biensen N.J. & Ford S.P. 1999. Novel insight into the control of litter size in pigs, using placental efficiency as a selection tool. Journal of Animal Science. 77: 1654-1658.
85 Wilson M.E., Biensen N.J., Youngs C.R. & Ford S.P. 1998. Development of Meishan and Yorkshire littermate conceptuses in either a Meishan or Yorkshire uterine environment to day 90 of gestation and to term. Biology of Reproduction. 58: 905-910.
86 Wilson M.E. & Ford S.P. 2001. Comparative aspects of placental efficiency. Journal of Reproduction & Fertility. 58 (Suppl): 223-232.
87 Wolf J., Záková E. & Groeneveld E. 2008. Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livestock Science. 115: 195–205.
88 Wu M.C., Hentzel M.D. & Dziuk P.J. 1988. Effect of stage of gestation, litter size and uterine space on the incidence of mummified fetuses in pigs. Journal of Animal Science. 66: 3202-3207.
89 Wu G., Bazer F.W., Cudd T.A., Meininger C.J. & Spencer T.E. 2004. Maternal nutrition and fetal development. Journal of Nutrition. 134: 2164-2172.
90 Wu G., Bazer F.W., Hu J., Johnson G.A. & Spencer T.E. 2005. Polyamine synthesis from proline in the developing porcine placenta. Biological Reproduction. 72: 842–850.
91 Wu G., Bazer F.W., Wallace J.M. & Spencer T.E. 2006. Board-invited review: intrauterine growth retardation: implications for the animal sciences. Journal of Animal Science. 84: 2316-2337.
93 Zhang S., Bidanel J.P., Burlot T., Legault C. & Naveau J. 2000. Genetic parameters and genetic trends in the Chinese x European Tiameslan composite line. Genetics Selection Evolution. 32: 41–56.
*publicado originalmente em:. Acta Scientiae Veterinariae. 37(Supl 1): s49-s60, 2009.
Tópicos relacionados
Autores:


Junte-se para comentar.
Uma vez que se junte ao Engormix, você poderá participar de todos os conteúdos e fóruns.
* Dados obrigatórios
Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.
Criar uma publicação




Você pode estar interessado em
.jpg&w=3840&q=75)