Respostas Morfofisiológicas de spp. ao Alagamento do Solo e a Síndrome da Morte do Capim-marandu Brachiaria
Publicado: 7 de novembro de 2011
Por: Moacyr Bernardino Dias-Filho
Introdução
"Em pastagens tropicais, o alagamento ou o encharcamento temporário do solo é relativamente comum. O excesso de água no solo da pastagem pode ser causado, naturalmente, por períodos chuvosos intensos, má drenagem do solo (e.g., solos com camada subsuperficial de impedimento) e a elevação sazonal do nível de rios e do lençol freático (DIAS-FILHO, 2005). Entretanto, o pisoteio do gado, o trânsito de máquinas, ou o impacto da chuva no solo descoberto, podem, paulatinamente, comprometer a capacidade natural de drenagem do solo por causa da compactação, tornando o solo da pastagem suscetível à ocorrência de períodos mais freqüentes e relativamente mais intensos de alagamento ou encharcamento. Assim, conforme discutido em Dias-Filho (2005), mesmo naqueles locais aonde o excesso de água no solo não seja naturalmente esperado, é possível que as práticas vigentes ou passadas de manejo contribuam para que, periodicamente, sejam instaladas condições, muitas vezes visualmente imperceptíveis, de excesso de água no solo.
O excesso de água no solo causa redução imediata na troca de gases entre a planta e o ambiente (ARMSTRONG et al. 1994; KOZLOWSKI, 2005; LIAO e LIN, 2001). A anoxia ou a hipoxia sofrida pelo sistema radicular altera o metabolismo celular, provocando queda imediata na respiração das raízes, tanto em plantas tolerantes como nas intolerantes (BRAENDLE e CRAWFORD, 1999; LIAO e LIN, 2001). A deficiência de oxigênio reduz a produção de ATP, afetando diversos aspectos do metabolismo celular (FUKAO e BAILEY-SERRES, 2004; SOUSA e SODEK, 2002). Essa queda na produção de ATP restringe o suprimento de energia para o crescimento das raízes, reduzindo o desenvolvimento geral da planta.
O problema da síndrome da morte do capim-marandu tem sido mencionado desde meados da década de 1990. Na Costa Rica, Zúñiga P. et al. (1998) associaram o problema a locais com umidade excessiva do solo e ao ataque de fungos. No Brasil, o problema da morte do capim-marandu também tem sido relacionado a pastagens formadas em solos mais úmidos (TEIXEIRA NETO et al. 2000; VALENTIM et al. 2000; DIAS-FILHO 2005; 2007; DIASFILHO e ANDRADE, 2005), principalmente em função da reconhecida baixa tolerância desse capim ao excesso de água no solo (DIAS-FILHO e CARVALHO, 2000). A lógica para a associação entre a baixa adaptação do capim-marandu ao encharcamento do solo e a síndrome da morte seria que o excesso de água no solo agiria como fator de predisposição para a instalação do problema. Essa predisposição estaria ligada a mudanças no comportamento da planta e nas características biológicas, físicas e químicas do solo, afetado pelo excesso de água.
Objetiva-se neste trabalho descrever o comportamento morfofisiológico de gramíneas tropicais e, particularmente, do capim-marandu (Brachiaria brizantha cv. Marandu), a hipoxia ou anoxia do sistema radicular. Discute-se, ainda, a possível associação entre o fenômeno da síndrome da morte do capim-marandu e as respostas do capim-marandu ao alagamento do solo.
Respostas morfofisiológicas de capins tropicais ao alagamento do soloEspécies comerciais
Poucos trabalhos têm sido publicados sobre o comportamento morfofisiológico de capins tropicais ao alagamento ou encharcamento do solo. Ensaios em que se avaliam a tolerância de gramíneas forrageiras ao alagamento do solo, por meio do estudo do comportamento morfofisiológico, geralmente necessitam ser conduzidos em ambiente controlado ou semicontrolado. Um problema metodológico comum a esses ensaios é que, quando não adequadamente planejados, o tempo de condução pode ultrapassar a capacidade dos vasos em manter o crescimento adequado das plantas do tratamento controle (não alagado). Isso se deve à rápida ocupação do volume de solo pelas raízes das gramíneas, ocasionando perda de vigor, em decorrência da queda na fertilidade do solo e à instalação de uma condição crônica de déficit hídrico. Esse fato leva a que, em algumas situações e para alguns parâmetros medidos, não seja possível detectar diferenças reais entre plantas alagadas e não alagadas, mesmo quando são comparadas espécies forrageiras, tidas como intolerantes ou pouco tolerantes ao alagamento do solo. Portanto, recomenda-se cuidado especial na interpretação de resultados oriundos desses ensaios.
Em trabalhos pioneiros de Baruch (1994a; 1994b) compararam-se respostas morfofisiológicas de quatro capins tropicais (Brachiaria mutica, Hyparrhenia rufa, Anogon gayanus e Echinochloa polystachya) sob alagamento do solo. Nesses trabalhos, constatou-se que sob alagamento, A. gayanus e H.
rufa fecharam rapidamente seus estômatos, reduzindo a condutância estomática e a fotossíntese líquida. A atividade da desidrogenase alcoólica (ADH) em A. gayanus sofreu aumento significativo sob alagamento, indicando maior sensibilidade daquela espécie a esse estresse (maior produção de etanol). No entanto, em B. mutica e E. polystachya, espécies típicas de ambientes alagados, a condutância estomática, a fotossíntese líquida e a atividade ADH não diferiram estatisticamente entre plantas alagadas e não alagadas. Segundo Baruch (1994a; 1994b), tanto B. mutica quanto E. polystachya apresentaram estruturas adaptativas ao alagamento, como o desenvolvimento de raízes adventícias e tecidos arenquimáticos, que permitiram melhor difusão de gases na planta.
Em um dos primeiros estudos publicados no Brasil, comparando a tolerância de diferentes espécies de Brachiaria ao alagamento do solo (DIAS-FILHO e CARVALHO, 2000), foi sugerido que a taxa diária de alongamento foliar poderia ser usada como parâmetro indicador precoce da tolerância de capins ao alagamento do solo. Sob alagamento, a taxa de alongamento foliar de B. brizantha cv. Marandu foi reduzida significativamente, enquanto que em B. decumbens e B. humidicola não foi possível detectar diferenças entre tratamentos. No capim-marandu, tal redução ocorreu a partir do 1º dia de alagamento do solo. No mesmo estudo, resultados apresentados em Dias-Filho (2005) mostram que o conteúdo de amido medido nas folhas, 6 dias após o início do alagamento, foi inversamente proporcional à tolerância relativa desses capins a esse estresse (Fig. 1), sugerindo que, sob alagamento do solo, as principais vias de metabolismo de açúcares foram alteradas nessa cultivar. A explicação para tal resposta seria que nas plantas mais suscetíveis ao alagamento, haveria menor demanda de carboidratos pelas raízes, causada pela diminuição no crescimento e metabolismo dessas estruturas. No entanto, o alagamento também reduziria o transporte de carboidratos das folhas para as raízes (LIAO e LIN, 2001). Todos esses eventos causariam acúmulo de fotoassimilados nas folhas, na forma de amido.
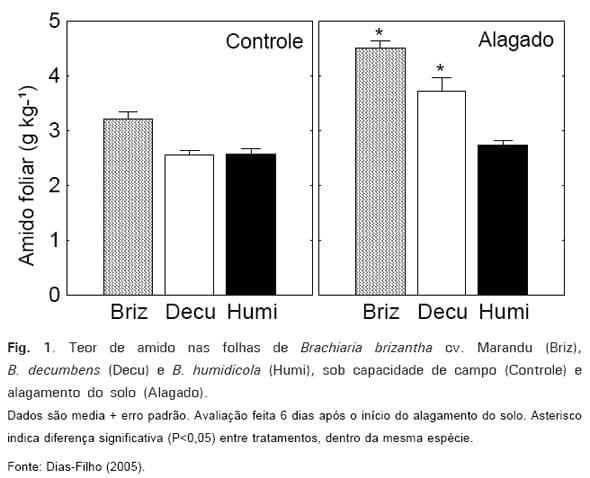
Outros parâmetros, medidos no decorrer do estudo, também indicaram a mesma tendência (Tabela 1), mostrando a baixa tolerância ao alagamento do solo do capim-marandu, a tolerância relativamente moderada de B. decumbens e a maior tolerância de B. humidicola.
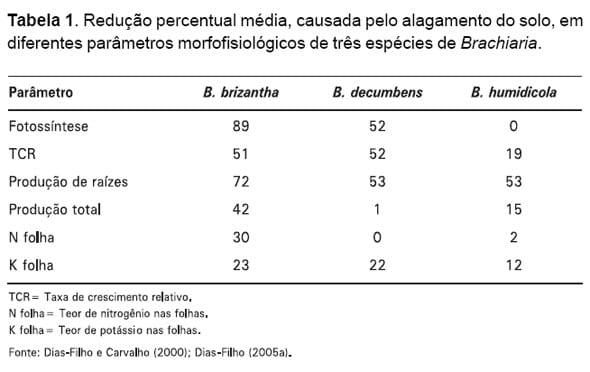
A forte queda observada na capacidade fotossintética do capim-marandu sob alagamento do solo (Tabela 1), poderia, pelo menos em parte, ser atribuída a fatores de natureza não estomática, como o acúmulo de amido nas folhas (Fig. 1), o qual provocaria inibição da fotossíntese (feedback inhibition) (LIAO e LIN, 2001). Entretanto, o teor de clorofila total também foi reduzido significativamente pelo alagamento do solo no capim-marandu, porém, não foi afetado nas demais espécies (DIAS-FILHO e CARVALHO, 2000), podendo ter contribuído para o comportamento fotossintético medido no ensaio (Tabela 1).
O acúmulo de açúcares solúveis em raízes de plantas sob alagamento do solo tem sido reportado como forma de tolerância a esse estresse (LIAO e LIN, 2001).
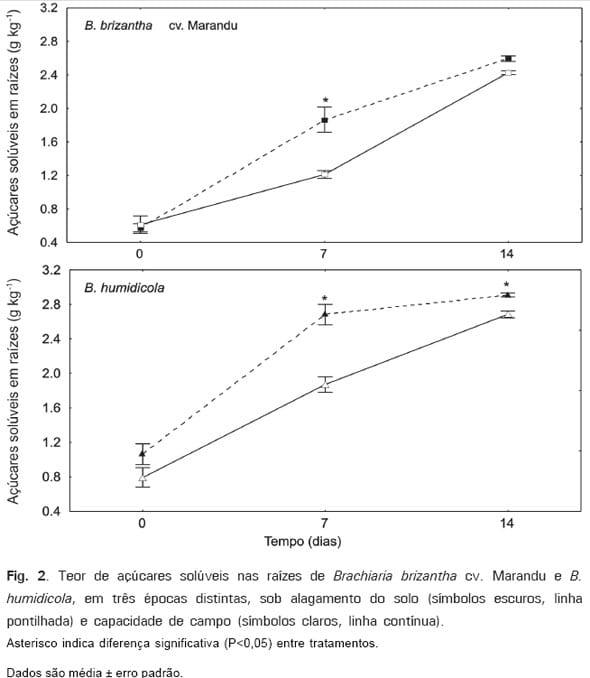
No entanto, em alguns estudos, têm-se constatado grande acúmulo de açúcares em raízes de plantas tidas como intolerantes ao alagamento do solo (CASTONGUAY et al. 1993; SU et al. 1998). Em estudo comparando respostas morfofisiológicas do capim-marandu e B. humidicola, capins com graus de tolerância opostos ao alagamento do solo, mostrou-se que, sob alagamento, ambas espécies acumularam mais carboidratos nas raízes, quando comparadas a plantas sob capacidade de campo. No entanto, esse acúmulo foi maior em B.
humidicola (Fig. 2). No mesmo estudo, observou-se acúmulo significativo de amido nas folhas do capim-marandu submetido a alagamento do solo (Fig. 1). O acúmulo de amido nas folhas e o maior conteúdo de açúcares solúveis nas raízes do capim-marandu sob alagamento do solo, aparentemente resultariam da diminuição da demanda por carboidratos por causa da redução do crescimento e das atividades metabólicas nas raízes. Esses resultados indicam que o acúmulo de amido nas folhas não resulta em deficiência de açúcares nas raízes do capimmarandu sob alagamento, e ainda que o nível de açúcares nas raízes não seria crítico para a tolerância dessa cultivar ao alagamento do solo.
Costa (2004) comparou o comportamento dos capins angola (Brachiaria mutica) e canarana verdadeira (Echinochloa polystachya), sob alagamento do solo. Embora esses capins sejam considerados tolerantes a esse estresse, constatou-se que, em ambas espécies, plantas alagadas sofreram reduções na área foliar, nas produções de massa seca de lâmina foliar e total, relação folha-haste e no número de perfilhos, quando comparadas a plantas cultivadas sob capacidade de campo. Isto é, mesmo em capins considerados tolerantes ao alagamento do solo, esse estresse pode causar diminuição no vigor e na capacidade produtiva da planta.
Acessos não comerciais de Brachiaria spp.
Em pesquisas desenvolvidas desde 1999, têm-se comparado a tolerância relativa ao alagamento do solo de novos acessos de capins do gênero Brachiaria, particularmente B. brizantha, visando a subsidiar programa de lançamento de novas cultivares de gramíneas forrageiras pela Embrapa (DIASFILHO, 2002; DIAS-FILHO e QUEIROZ, 2003; MODESTO, 2005). Nesses estudos, acessos pré-selecionados de Brachiaria spp., oriundos da Embrapa Gado de Corte e ainda não lançados no mercado como cultivares comerciais, são comparados com a cultivar Marandu ou Xaraés, visando classificar a tolerância relativa desses acessos ao alagamento do solo. Os resultados obtidos até o momento indicam que grande parte dos acessos já testados possui tolerância ao alagamento do solo superior à do capim-marandu. Esses resultados confirmam que pode existir grande variabilidade, mesmo dentro da mesma espécie, na capacidade de adaptação das plantas ao alagamento do solo. Embora variações na tolerância ao alagamento do solo normalmente sejam baseadas em adaptações anatômicas e bioquímicas complexas, no presente caso, onde a maioria dos acessos testados é morfologicamente semelhante, a variação no grau de tolerância poderia estar principalmente baseada em diferenças ligadas a adaptações metabólicas desses acessos.
Estudo comparando o comportamento morfofisiológico de seis acessos de B.
brizantha sob alagamento do solo mostrou que os acessos diferiram quanto à tolerância relativa a esse estresse (DIAS-FILHO e QUEIROZ, 2003). Apesar da tolerância diferenciada entre acessos, a produção de raízes foi profundamente afetada (>50%) em todos eles, mesmo naqueles classificados como relativamente mais tolerantes (Fig. 3). Em estudo semelhante, onde foi comparada a tolerância relativa ao alagamento de cinco acessos de B.
brizantha (DIAS-FILHO, 2002), a produção de raízes também foi um dos parâmetros morfofisiológicos mais afetados pelo alagamento do solo (Fig. 4).
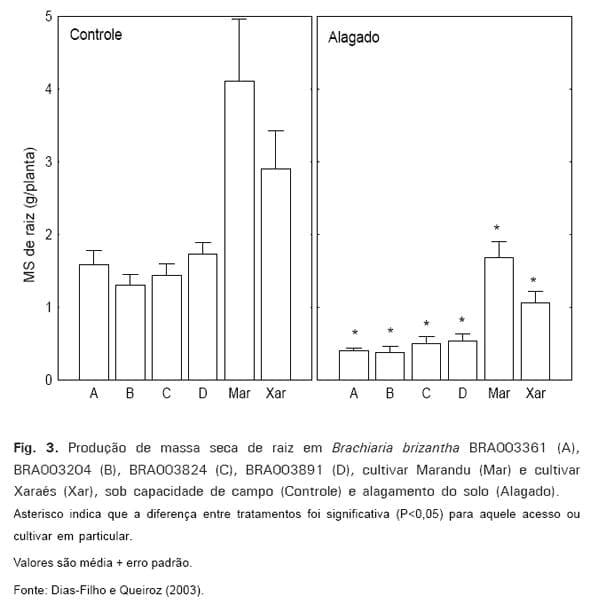
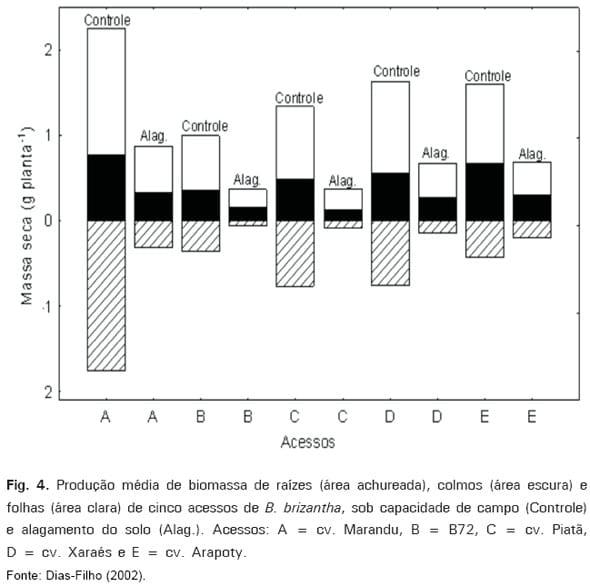
O fato das raízes serem os órgãos mais afetados pelo alagamento do solo traria conseqüências particularmente drásticas para capins menos tolerantes a esse estresse. A razão para isso seria que as pastagens normalmente estão sujeitas ao encharcamento temporário ou ao alagamento do solo durante a estação chuvosa, período no qual as atividades fisiológicas são máximas na planta. Tal fato intensificaria os efeitos nocivos do alagamento do solo nas plantas menos tolerantes, prejudicando, ainda, a criação de condições (e.g., construção de sistema radicular mais vigoroso) que permitiriam maior tolerância a estresses, durante a estação seca subseqüente (e.g., déficit hídrico ou o superpastejo).
Dessa forma, pastagens formadas com capins pouco tolerantes ao excesso de água no solo, que fossem submetidas a situações crônicas de anoxia ou hipoxia do sistema radicular, estariam mais suscetíveis à degradação por causa da perda de vigor, a alterações metabólicas e, conseqüentemente, da capacidade em responder eficientemente a estresses adicionais (DIAS- FILHO, 2005; DIAS-FILHO, 2007).
Espera-se que com a evolução das pesquisas com esses novos acessos, em futuro próximo, seja possível lançar no mercado cultivares de B. brizantha com as mesmas características positivas que impulsionaram o uso do capimmarandu no Brasil, porém, sem a sua baixa tolerância ao alagamento do solo.
Essa baixa tolerância a condições periódicas de encharcamento do solo, aparentemente, seria a principal causa dessa cultivar estar sendo paulatinamente inviabilizada em alguns locais do Trópico Úmido.
Síndrome da morte do capimmarandu e o alagamento do solo
Em meados dos anos de 1990, começaram a surgir relatos de áreas de pastagens de Brachiaria brizantha cv. Marandu com sintomas de murchamento e morte, no Acre, Pará e em Rondônia (TEIXEIRA NETO et al.
2000; VALENTIM et al. 2000). Em 1998, já existiam vastas áreas de pastagem de capim-marandu, na Amazônia Legal, com sintomas semelhantes (TEIXEIRA NETO et al. 2000; VALENTIM et al. 2000). Relatos anteriores aos reportados no Brasil, dão conta que em 1997, já havia ocorrência de morte do capim-marandu em áreas sujeitas ao encharcamento temporário do solo no Trópico Úmido da Costa Rica (ZÚÑIGA P. et al. 1998). Ao conjunto de fatores que causaram as alterações nas pastagens, denominou-se síndrome da morte de pastagens do capim-marandu (TEIXEIRA NETO et al. 2000; VALENTIM et al. 2000).
Essa síndrome manifesta-se durante a época chuvosa, principalmente em áreas que apresentam solos de baixa permeabilidade. Estudos realizados no Acre (VALENTIM et al. 2000; ANDRADE et al. 2003), no Pará (DIAS FILHO e CARVALHO, 2000; DIAS-FILHO, 2002; TEIXEIRA NETO et al. 2000) e na Costa Rica (ZÚÑIGA P. et al. 1998), sugeriram que a mortalidade do capimmarandu, provavelmente, se deveria à associação da falta de adaptação dessa cultivar ao encharcamento periódico do solo com o ataque de fungos, que são favorecidos pela condição de saturação de água no solo e a conseqüente diminuição de vigor do capim. Na Costa Rica, Zúñiga P. et al.
(1998) isolaram estirpes de Fusarium, Rhizoctonia, e Pythium em pastagens de capim-marandu, onde o problema havia sido detectado. Testes de patogenicidade desses fungos em genótipos de B. brizantha, incluindo as cultivares Marandu e Xaraés, e em B. dictyoneura, sob encharcamento do solo, confirmaram a alta susceptibilidade do capim-marandu a estes patógenos. Com o solo na capacidade de campo, apenas sintomas leves foram constatados. A cultivar Xaraés apresentou tolerância relativamente maior e a B. dictyoneura não foi afetada.
A alta correlação entre o fenômeno da síndrome da morte do capim-marandu e a drenagem deficiente do solo fica bastante evidente no Estado do Acre.
Nesse estado, a síndrome da morte do capim-marandu já é a principal causa de degradação das pastagens (DIAS-FILHO e ANDRADE, 2005), sendo que a maioria dos solos aonde tais pastagens foram formadas, apresenta algum tipo de deficiência na capacidade de drenagem (VALENTIM et al. 2000).
Inferir que o problema da morte do capim-marandu estaria simplesmente relacionado à baixa adaptação dessa cultivar ao alagamento do solo, poderia ser considerada uma concepção superficial desse fenômeno. Certamente, a baixa tolerância do capim-marandu ao alagamento do solo, por si só, tornaria a planta mais suscetível a estresses bióticos, como o ataque de patógenos.
No entanto, alterações metabólicas causadas pelo alagamento do solo no capim-marandu, provavelmente seriam a principal causa que tornaria essa cultivar mais suscetível ao ataque de patógenos. Por exemplo, a descoberta de que o capim-marandu tende a acumular amido nas folhas e açúcares solúveis nas raízes, quando sob alagamento do solo (Fig. 1 e 2), indica que o metabolismo de açúcares estaria sendo afetado por tal estresse nessa cultivar. Como mudanças no metabolismo de açúcares têm sido observadas em plantas infectadas por patógenos, e vários genes envolvidos nas respostas a estresse e defesa parecem responder ao metabolismo de açúcares da planta (TADEGE et al. 1999), seria plausível inferir que, sob alagamento do solo, o capim-marandu poderia sofrer alterações em seus mecanismos bioquímicos de defesa, tornando-se suscetível aos agentes bióticos que, em condições normais, não seriam capazes de provocar danos com a mesma intensidade nessa cultivar. Em outras palavras, a exposição dessa cultivar ao alagamento do solo, poderia estar provocando disfunção nos mecanismos metabólicos de defesa da planta.
De fato, estudos conduzidos com diferentes espécies vegetais têm mostrado estreita relação entre a anoxia do sistema radicular e o aumento na suscetibilidade aos danos causados por infecções de patógenos (BURGESS et al. 1998; 1999; ROBIN et al. 2001). Os mecanismos sugeridos para essa relação estariam ligados a modificações metabólicas que dificultariam o reconhecimento do patógeno pela planta ou que facilitariam a sua colonização e desenvolvimento. Por exemplo, em tomateiro cultivado sob deficiência de oxigênio nas raízes e inoculado com Pythium, foram encontrados sintomas típicos de ataque desse patógeno, enquanto que as plantas sem deficiência de oxigênio permaneciam saudáveis (CHÉRIF et al.
1997). Segundo Chérif et al. (1997), aumentos na atividade das lipoxigenases, isoenzimas ligadas à proteção da planta contra o ataque de agentes bióticos como fungos (PORTA e ROCHA-SOSA, 2002), observados nas raízes das plantas sob deficiência de oxigênio, poderiam levar à degradação e à desorganização dos lipídeos da membrana celular, favorecendo a colonização das raízes por patógenos.
Outro mecanismo relacionado à maior sensibilidade a infecções por patógenos, em plantas cultivadas em solos com excesso de umidade, seria a capacidade em atrair patógenos exercida por compostos orgânicos solúveis, exudados pelas raízes (BRAENDLE e CRAWFORD, 1999). Por exemplo, a exudação de etanol (composto metabolizado pela planta sob anoxia), normalmente em maior abundância nas espécies menos tolerantes ao alagamento (BARTA, 1987), teria a capacidade de atrair zoósporos de fungos e fornecer substratos para a colonização de micélios no tecido vegetal (ALLEN, 1974; YOUNG e NEWHOOK, 1977). Ademais, solos alagados teriam, ainda, a capacidade de aumentar a mobilidade de zoósporos, facilitando a intensidade e o alastramento do problema.
Finalmente, estresses adicionais, como os baixos níveis de fósforo e potássio no solo e o superpastejo, em conseqüência da influência que teriam no comportamento morfofisiológico da planta, poderiam agir sinergeticamente para potencializar os efeitos causados pelo problema da síndrome da morte do capim-marandu.
Considerações Finais
O problema da síndrome da morte do capim-marandu vem causando sérios problemas para a produtividade da pecuária, já sendo importante causa de degradação de pastagens em determinados locais da Amazônia Brasileira.
Evidências obtidas experimentalmente, ou com base em observações de campo, sugerem forte correlação entre o aparecimento do problema e solos com drenagem deficiente, situados em regiões com períodos chuvosos intensos, com altas temperaturas e altos níveis de umidade do ar.
Por causa da baixa tolerância do capim-marandu a condições persistentes de encharcamento do solo, e a alterações morfofisiológicas sofridas pela planta sob esse estresse, seria possível inferir que o excesso de água no solo atuaria como fator de predisposição para a instalação do problema da síndrome da morte nessa cultivar. Plantas afetadas pelo excesso hídrico seriam, possivelmente, mais suscetíveis a danos causados por infecções de patógenos ou outros agentes bióticos, além de, naturalmente, terem a produtividade bastante diminuída por esse estresse.
Dados experimentais e evidências da literatura científica mostram que determinados aspectos do comportamento fisiológico e morfológico do capimmarandu e de outras espécies vegetais, sob alagamento do solo, como alterações fisiológicas (e.g., metabolismo de açúcares) ou morfológicas (e.g., diminuição no vigor e crescimento do sistema radicular), poderiam aumentar a predisposição a infecções, ou mesmo causar regeneração insuficiente das raízes já infectadas por patógenos ou outros agentes bióticos. Além disso, com base no comportamento de outras espécies vegetais, sabe-se que sob alagamento do solo, compostos orgânicos solúveis (e.g., etanol) podem ser exudados pelas raízes, tendo a capacidade de atrair zoósporos de fungos e fornecer substratos para a colonização de micélios no tecido vegetal.
Cerca de 10 anos após os primeiros relatos da sua ocorrência no Brasil, ainda não existe entendimento amplo sobre o fenômeno da síndrome da morte do capimmarandu. Dessa forma, com base no que já foi estudado sobre o assunto, acreditase que as ações que visem ao entendimento desse problema devam, a partir de agora, basear-se em estudos de natureza fisiológica e bioquímica que busquem compreender a influência exercida por agentes bióticos no desempenho do capimmarandu, cultivado sob uma combinação de estresses abióticos, como a anoxia do sistema radicular, baixos níveis de P e K no solo e o superpastejo.
Tópicos relacionados:
Autores:

Embrapa
Recomendar
Comentário
Compartilhar

Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.
