Competição e Sucessão Vegetal em Pastagens
Publicado: 1 de dezembro de 2011
Por: Moacyr Bernardino Dias-Filho (Engenheiro Agrônomo - Embrapa Amzônia Oriental)
Introdução
Em comunidades vegetais, as plantas irão afetar-se mutuamente através da competição. A competição resulta da reação de uma planta sobre os fatores (recursos) ambientais físicos assim como o efeito desses fatores modificados nos competidores. A competição só passará a existir se os recursos disponíveis no ambiente (luz, água do solo, nível de nutriente, espaço físico) não estiverem em excesso as necessidades das plantas. A competição tem início quando a disponibilidade de um único recurso cai abaixo das necessidades conjuntas das plantas (Donald, 1963). O espaço físico é o único recurso comum pelo qual todas as espécies competem em conjunto. A necessidade comum por luz, água e nutrientes, no entanto, significa que a competição por vários fatores é inevitável, apesar da quantidade de recursos requerida por cada espécie poder variar (Turkington & Mehrhoff, 1990).
Ecossistemas de pastagens cultivadas e nativas diferem em complexidade de outros ecossistemas agrícolas nos quais, geralmente, apenas a cultura e as plantas daninhas seriam os principais componentes do sistema. Em pastagens, a presença do animal, como o “agente de colheita” da cultura amplia, em muito, a complexidade do sistema, interferindo direta e indiretamente nos padrões de competição e sucessão.
A competição é um importante componente que controla os processos que levam ao declínio em produtividade, perda de espécies desejáveis e a invasão de plantas daninhas, assim como aqueles associados com o aumento da produtividade e estabilidade da pastagem (Tow & Lanzeby, 2001). A natureza dessa competição ainda não é completamente entendida. Existem outros fatores, abióticos e bióticos, os quais podem ter um profundo efeito nas condições em que a competição ocorre.
Em pastagens, a competição poderá ocorrer desde o momento da semeadura, quando a(s) forrageira(s) recém-germinada(s) têm que competir por espaço, luz, água e nutrientes com outros indivíduos da mesma espécie, ou com outras espécies, deliberadamente plantadas pelo produtor, ou aquelas que estavam presentes no banco de sementes do solo. Essas últimas seriam basicamente as “espécies colonizadoras” (sucessionais), as quais teriam o papel de promover o processo de sucessão vegetal na pastagem. O objetivo principal das práticas de manejo, desde o estabelecimento da pastagem, é otimizar a produção de forma sustentável. Assim, essas práticas de manejo (adubação, sistema de pastejo, controle de plantas invasoras etc.), juntamente com as condições ambientais, terão grande impacto na capacidade competitiva de cada espécie presente no sistema e, conseqüentemente, no padrão de sucessão vegetal que se desenvolverá na área.
Os princípios da competição e da sucessão vegetal são diretamente relevantes ao desafio de manter uma composição botânica ideal na pastagem, livre de plantas daninhas (invasoras), e que permita a obtenção de altos níveis de produtividade e utilização. A composição botânica é influenciada pelas relações competitivas, podendo ser indicadora de um estádio na sucessão vegetal e, conseqüentemente, da produtividade da pastagem. Assim, o processo de degradação de pastagens estaria intimamente relacionado com os padrões de competição e o processo de sucessão que se desenvolve na pastagem. Conforme proposto em Dias-Filho (2003), uma pastagem poderia ser considerada degradada ou em degradação dentro de dois extremos de composição botânica (Figura 1). Em um extremo, a degradação pode ser inicialmente caracterizada apenas pela mudança na composição botânica da pastagem, em decorrência do aumento na proporção de plantas daninhas (invasoras) e da diminuição da proporção de capim. Nessa situação, a degradação da pastagem poderia ser denominada “degradação agrícola”, isto é, a capacidade da pastagem para produzir economicamente (do ponto de vista agropecuário), estaria temporariamente diminuída ou inviabilizada, devido à pressão competitiva exercida pelas plantas daninhas sobre o capim, causando, portanto, queda acentuada na capacidade de suporte da pastagem.
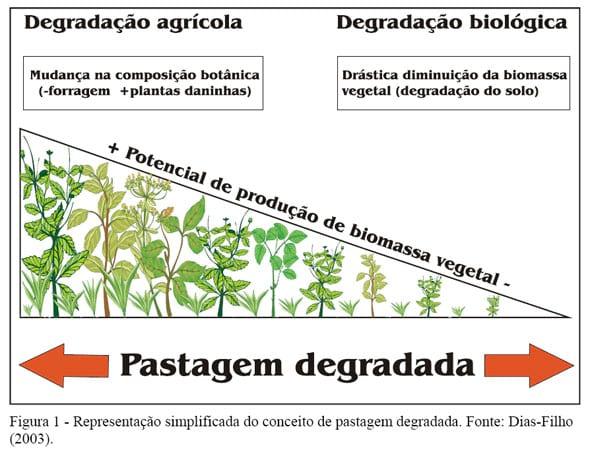
Em outro extremo, a degradação da pastagem pode ser caracterizada pela drástica diminuição da biomassa vegetal da área, provocada pela degradação do solo, que, por diversas razões de natureza química (mineração dos nutrientes e acidificação), física (erosão e compactação) ou biológica (perda da matéria orgânica), estaria perdendo a capacidade de sustentar produção vegetal significativa. Nessa condição mais drástica de degradação, a gramínea forrageira seria gradualmente substituída por espécies pouco exigentes em fertilidade do solo e de menor valor nutritivo e capacidade de produção de biomassa vegetal adaptadas a essas condições, ou, simplesmente, seria substituída por áreas de solo descoberto, altamente vulneráveis à erosão. Nesse outro extremo, a degradação poderia ser denominada “degradação biológica”, pois a capacidade da área em sustentar a produção vegetal estaria comprometida. Nessa situação extrema de produtividade da pastagem, houve “reversão” no processo de sucessão vegetal para o estádio de solo nu, via de regra causada por práticas inadequadas de manejo, como altas pressões de pastejo, uso excessivo do fogo etc., as quais provocam danos severos à parte aérea e subterrânea da vegetação, erosão do solo e queda acentuada da fertilidade e da matéria orgânica.
O uso de práticas adequadas de manejo visando à manutenção ou recuperação da produtividade depende do entendimento apropriado dos princípios envolvidos na competição e sucessão vegetal. Assim, as práticas agronômicas empregadas nessas atividades devem ser baseadas, também, em conceitos ecológicos.
Definições e Conceitos
Silvertown & Wilson (2000) descrevem competição como a interação entre plantas vizinhas na qual cada indivíduo suprime a performance do outro.
Grime (1979; 2001) define competição como “a tendência de plantas vizinhas em utilizar o mesmo quantum de luz, íon de nutriente mineral, molécula de água, ou volume de espaço”. De acordo com essa definição, a competição se refere exclusivamente à captura de recursos. A teoria de Grime postula que existem três estratégias básicas nas plantas: competidora, tolerante ao estresse e ruderal. A competitividade, isto é, a habilidade em dominar uma comunidade vegetal, não seria determinada por uma única característica; mas sim por uma “síndrome” de características, as quais incluem taxa máxima de crescimento relativo, taxa de assimilação líquida e razão de área foliar. Ele classifica os bons competidores em três categorias principais: (1) plantas altamente competitivas (C), as quais podem usufruir as condições ambientais favoráveis, absorvendo rapidamente nutrientes e crescendo com rapidez, dominando, conseqüentemente, a vegetação; (2) Plantas tolerantes ao estresse (S), que são adaptadas, ou pelo menos tolerantes as condições de escassez de recursos; e (3) espécies ruderais (R) que têm a habilidade para invadir e se desenvolver sob condições características de áreas alteradas. De acordo com Grime (1979) a planta será competitivamente superior se for capaz de capturar os recursos com mais rapidez do que as plantas vizinhas. Isso pode estar relacionado a características particulares das plantas, como alta taxa de crescimento relativo.
Por outro lado, Tilman (1982) postula que a habilidade da planta em continuar a extrair recursos mesmo em níveis bem baixos deste recurso seria importante indicador da capacidade competitiva dessa espécie. Isto é, um bom competidor é capaz de desempenhar bem apesar da escassez de recursos. Tilman baseou sua definição de habilidade competitiva na teoria que, através de alguns anos, o competidor de sucesso é a espécie (dentre aquelas inicialmente presentes) que tem a capacidade de reduzir a concentração do recurso edáfico limitante (ex. N) ao nível mais baixo e mesmo assim manter a sua população, isto é, é a espécie com o requerimento de recursos mais baixo. Com relação a pastagens, a teoria de Tilman é mais adequada para pastagens pouco pastejadas e que recebem poucos insumos, do que para aquelas pastejadas intensamente e que recebem regularmente altas quantidades de nutrientes.
Existe certo consenso quanto ao fato que a teoria de Grime aplica-se adequadamente a ambientes produtivos, enquanto que a teoria de Tilman pode ser mais apropriada para o entendimento de sistemas com baixos níveis de nutrientes e através de maior espaço de tempo.
Em resumo, na teoria proposta por Grime, planta competitiva é aquela que possui elevada velocidade de utilização dos recursos do meio, tornando esses recursos indisponíveis para as plantas vizinhas. Assim, uma planta competidora apresenta elevada taxa de crescimento relativo (TCR). Por outro lado, a teoria estabelecida por Tilman postula que plantas competitivas necessitam de menos recursos e, portanto, conseguem sobreviver em ambientes desfavoráveis, ou seja, com baixo aporte de água, nutrientes e/ou luz.
Características Morfofisiológicas que Afetam a Competitividade das Plantas
Muitos dos atributos que caracterizam uma espécie vegetal como tolerante ao estresse devem também caracterizar essa espécie como sendo competidora eficaz em tais habitats (Peltzer & Wilson, 2001). De acordo com Grime (2001), a capacidade competitiva da planta estaria relacionada a uma combinação de atributos que facilitariam a interceptação de recursos. Grime (2001) alerta, no entanto, que essas características não seriam por si só diagnósticos da capacidade competitiva. Alguns desses atributos, que seriam importantes em ecossistema de pastagem, são descritos a seguir:
Órgãos de reserva
A mobilização de reservas de energia acumuladas em órgãos subterrâneos permite a formação extremamente rápida do dossel. A ocupação de espaço no dossel seria a vantagem competitiva proveniente dessa característica.
Altura
Segundo Grime (2001), em situações onde a vegetação herbácea seja densa (como em pastagens), pequenas diferenças em altura podem significar grandes mudanças na intensidade, direção e qualidade da radiação. A habilidade da plântula ou planta em estabelecimento competir por luz pode depender da capacidade de suas folhas alcançarem posições superiores no dossel. O crescimento em altura dependeria, principalmente, do suprimento de energia proveniente de órgãos de reserva e da capacidade fotossintética atual e também, da morfologia da planta. Em pastagens cultivadas tropicais, como aquelas encontradas na região amazônica, grande proporção das invasoras lenhosas apresenta a capacidade de crescimento rápido em altura, tornando-se plantas altamente competitivas com as gramíneas da pastagem (Dias Filho, 1990; 2003).
Visando a predizer coeficientes de competição para consórcios de plantas Freckleton & Watkinson (2001) estudaram dados de trabalhos publicados sobre interações competitivas e encontraram que o atributo que melhor se correlacionou com o coeficiente de competição é a diferença no tamanho máximo das espécies. Em revisão feita por Nurjaya & Tow (2001) entre as características competitivas de diferentes genótipos, destaca-se que atributos relacionados ao tamanho da planta e a características foliares, como a razão de área foliar (RAF) e, principalmente, a área foliar específica (AFE, área foliar:massa foliar) estariam significativamente correlacionados com a capacidade competitiva. Outros estudos também têm ressaltado a relação positiva entre a capacidade competitiva e a altura (tamanho) da planta (por exemplo, Mennan & Isik 2004), em outros, porém, esta relação não tem se mostrado tão evidente (Juskiw et al. 2000).
Expansão lateral
Em pastagens, algumas plantas invasoras lenhosas podem atingir alturas relativamente elevadas sem, no entanto, conseguir captar grande proporção dos recursos disponíveis no ambiente, isto é, sem afetar, de forma mais intensa, a disponibilidade de recursos para as plantas vizinhas. Assim, espécies com maior capacidade competitiva seriam aquelas nas quais o crescimento em altura estaria associado à expansão lateral que resultasse em alta densidade de ramos e raízes.
Estudos têm mostrado alta correlação entre a biomassa radicular e a capacidade competitiva (Tilman & Wedin, 1991; Cahill & Casper, 2000). Em estudo comparando características de raízes finas de 11 espécies de árvores, Comas & Eissenstat (2004) encontraram que as raízes das espécies de crescimento mais rápido geralmente apresentam maior comprimento específico, menor diâmetro, maior grau de expansão, e menor concentração de fenóis do que daquelas de crescimento mais lento, sugerindo diferenças nas estratégias de exploração do solo e defesa das raízes. Estas características teriam reflexo direto na capacidade competitiva dessas espécies.
Taxa de crescimento
Para que a planta alcance, com eficiência, rápido crescimento em altura, expansão de raízes e folhas e acúmulo de reservas, é necessário ter rápida taxa de crescimento. Isto é, a planta será competitivamente superior se tiver capacidade de capturar os recursos com maior rapidez do que as plantas vizinhas.
Nutrientes foliares
A aptidão em acumular nutrientes (principalmente nitrogênio e fósforo) nas folhas, geralmente está relacionada a altas taxas de crescimento, as quais seriam conseqüência da maior capacidade fotossintética. Espécies de crescimento mais lento (menor capacidade competitiva) normalmente acumulariam menores proporções de nitrogênio e fósforo nas folhas.
Área foliar específica (AFE)
Em geral a AFE (área foliar/massa foliar) está positivamente relacionada com a taxa de crescimento relativo. O investimento em estruturas de defesa (por exemplo, anti-herbivoria), como compostos fenólicos, e o acúmulo de parede celular reduzem a AFE, aumentam a longevidade foliar e diminuem o conteúdo de nitrogênio da folha (Lambers & Poorter, 1992).
Espécies que apresentam valores mais altos de AFE, geralmente também apresentam altas concentrações de nitrogênio, altas taxas fotossintéticas por unidade de N foliar (revisado por Lambers & Poorter, 1992). Por outro lado, as espécies com valores baixos de AFE, geralmente têm maiores concentrações de compostos secundários e parede celular e maior longevidade de folhas e raízes (Choong et al. 1992, Ryser 1996, Reich 1998). Assim, espécies com altos valores de AFE seriam mais eficientes na captura de recursos, enquanto que aquelas com baixos valores de AFE teriam características que favoreceriam a conservação dos recursos capturados. Normalmente, espécies de ambientes mais produtivos têm maior AFE do que aquelas características de ambientes menos produtivos (Poorter & de Jong 1999).
Eficiência na busca e uso de recursos
A eficiência da planta em forragear por recursos tem grande importância na sua capacidade competitiva (revisado por Grime, 2001). Essa eficiência está diretamente correlacionada com a plasticidade da espécie em ser capaz de rapidamente responder a variações na disponibilidade de recursos no ambiente através da proliferação de raízes finas, expansão da área foliar e crescimento de ramos e pecíolos. Assim, rápidos ajustes morfológicos em resposta a mudanças na disponibilidade de recursos facilitariam a captura de água, luz, nutrientes e espaço pela planta, tornando-a mais competitiva (Rubio et al. 2003; Casper & Jackson, 1997; Fransen et al. 2001; Aikio & Markkola, 2002; Rees & Bergelson, 1997; Herben & Suzuki, 2002; Novoplansky, 2003; Oborny et al. 2000).
Resposta ao dano
Em ambiente de pastagem, onde a vegetação está sujeita, com certa freqüência, a desfolha, através do pastejo, da roçagem, ou da queima, os competidores mais eficientes são aqueles que têm a habilidade em responder a desfolha por meio do rápido restabelecimento do dossel através da promoção de recrescimento das folhas danificadas ou através da expansão de novos ramos. Nurjaya & Tow (2001) também sugerem que a aptidão para se recuperar da desfolha seria importante no desempenho competitivo de espécies vegetais em ecossistema de pastagem.
Palatabilidade
Plantas com crescimento rápido e oriundas de habitats produtivos, via de regra têm folhagem relativamente mais palatável, isto é, sem a presença ou com baixos níveis de compostos químicos que conferem defesa contra a herbivoria, enquanto que plantas com restrição de recursos e menor crescimento teriam maior quantidade de compostos secundários tóxicos a herbívoros e patógenos (Almeida-Cortez et al. 2004a; 2004b; Coley et al. 1985). Baixo investimento dos recursos capturados pela planta em mecanismos de defesa anti-herbivoria normalmente tornam essa planta mais competitiva (revisado por Grime, 2001). Segundo Grime (2001), espécies ou genótipos que utilizam grande proporção de seus recursos em defesas químicas e físicas podem tornar-se vulneráveis a exclusão por competidores que alocam seus recursos preferencialmente para a produção de novas raízes e folhas.
Segundo Nurjaya & Tow (2001), em ambiente de pastagem, a importância relativa ou absoluta de determinada característica morfofisiológica na capacidade competitiva estaria dependente dos seguintes fatores: a) estádio de desenvolvimento da planta; b) densidade populacional; c) eficiência no armazenamento de reservas de minerais e carboidratos nos tecidos para uso em situações de estresse; e d) plasticidade nas respostas morfofisiológicas a mudanças ambientais, desfolha devido a pastejo e nível de nutriente no solo.
A correlação positiva entre a capacidade competitiva e a plasticidade fisiológica pôde ser verificada em experimento desenvolvido por Poorter & Lambers (1986). Nesse experimento, dois genótipos de Plantago major com diferentes graus de plasticidade foram cultivados em ambiente controlado com diferentes freqüências ou flutuações no nível de nutrientes do solo. Quando cultivados conjuntamente e sob maiores freqüências na flutuação dos níveis de nutrientes do solo, o genótipo com a maior plasticidade fisiológica cresceu mais rápido, enquanto que o genótipo de menor plasticidade teve o seu crescimento reduzido.
Relação entre Competição e a Dinâmica de Invasão de Pastagens
A invasão de pastagens por espécies lenhosas é um evento comum tanto em pastagens nativas quanto cultivadas (Dias Filho, 2003). A erradicação de plantas invasoras lenhosas de pastagens normalmente é tarefa difícil de ser concretizada, retratando, de certa forma, a nossa limitação de conhecimento sobre as interações entre gramíneas e árvores.
Em ecossistema de pastagem (nativas e cultivadas), assim como em outros ecossistemas, a competição não está restrita a uma única situação de produtividade, mas sim dentro de um largo espectro de situações de produtividade (Wilson 1991, Reader et al. 1993, Wilson 1998). Em geral, com o aumento da produtividade a competição sofre uma inversão, passando a ser mais intensa acima (luz) do que abaixo (água e nutrientes) do solo (Wilson 1993a; 1993b; Wilson & Tilman 1993; 1995). Dessa forma, a invasão de espécies lenhosas em pastagens pode estar relacionada a mudanças na disponibilidade de recursos para as raízes e parte aérea.
Os efeitos competitivos, isto é, o grau no qual as plantas superam as plantas adjacentes, normalmente aumentam com o tamanho da planta (Grace 2001), sendo maior em comunidades mais produtivas (Wilson 1999). O sucesso das plantas lenhosas de maior porte em suprimir as gramíneas em áreas de pastagens pode estar relacionado ao fato de que as árvores são capazes de acumular mais biomassa.
De acordo com Reynolds & Pacala (1993) e Schwinning & Weiner (1998), existe grande diferença no fundamento da competição por recursos aéreos (luz) e subterrâneos (água e nutrientes). Isto se deve ao fato da luz ser suprida ao longo de uma dimensão espacial, enquanto que os nutrientes e a água seriam geralmente supridos de uma forma tridimensional.
Assim, de acordo com os padrões de alocação de carbono, as diferentes formas de crescimento (isto é, árvores e gramíneas) teriam eficiências diferenciadas na competição por luz ou por água e nutrientes. As árvores e arbustos por terem padrão de alocação relativa de biomassa para as partes aéreas, traduzidas em maior altura, seriam melhores competidoras de luz, diminuindo a disponibilidade desse recurso para as gramíneas (com estatura mais baixa). As gramíneas, por sua vez, com suas raízes fibrosas e abundantes, podem ser mais eficientes em esgotar os recursos do solo devido a maior abundancia relativa de raízes, em comparação às árvores. Dessa forma, o processo de invasão de pastagens por árvores ocorreria não somente devido ao maior volume individual de biomassa das árvores, mas também em função do padrão de alocação de carbono das árvores permitir que compitam eficientemente por luz, causando diminuição desse recurso para as gramíneas vizinhas, e que usem relativamente menos nutrientes. Assim, os padrões de biomassa contribuiriam para a habilidade competitiva em situações em que os competidores fossem suficientemente diferentes. Então, plântulas de espécies lenhosas teriam inicialmente pequena vantagem competitiva sobre gramíneas. No entanto, à medida que essas plantas aumentassem em massa e altura, elas se tornariam mais competitivas.
Diversas plantas daninhas de pastagens são espécies invasoras, geralmente introduzidas de outras regiões. Sabe-se que muitas plantas invasoras mostram-se mais vigorosas no local de introdução do que no ambiente nativo (Crawley 1987; Blossey & Nötzold, 1995; Mack et al. 2000). Uma explicação seria que essas espécies encontram na área invadida ambiente livre de inimigos naturais, principalmente herbívoros específicos. Esta condição pode influenciar o vigor da planta de duas formas. Os recursos que são normalmente perdidos para inimigos ou na produção de defesas podem ser alocados para o crescimento ou reprodução, mediante resposta fenotípica plástica (“Hipótese da liberação do inimigo”, Bazzaz et al., 1987; Tilman, 1999; Keane & Crawley, 2002). Alternativamente, plantas introduzidas que usufruem um longo período de tempo livre de inimigos naturais (por exemplo: insetos herbívoros), podem abdicar estratégias de defesa anti-herbivoria, permitindo que essas plantas realoquem os recursos previamente gastos em defesa, para características que aumentem a capacidade competitiva (“Hipótese da evolução do aumento da capacidade competitiva” proposta por Blossey & Nötzold, 1995). É possível supor que o sucesso de muitas plantas invasoras exóticas de pastagens no Brasil pode ser parcialmente explicado por essas teorias. Experimentos desenvolvidos no hemisfério Norte tanto têm confirmado (por exemplo, Siemann & Rogers, 2003) como refutado (Vilà et al. 2003) a hipótese da evolução da capacidade competitiva.
Impactos do manejo sobre a competição e a sucessão
As plantas forrageiras que compõem o ecossistema de pastagem devem ser capazes de adaptar-se a mudanças no ambiente e serem eficientes na captura de recursos em situações de competição a fim de desempenharem papel importante na composição botânica da pastagem. Dentre os atributos necessários para o sucesso da espécie, a plasticidade fenotípica, isto é, a capacidade de ajustar suas características morfológicas e fisiológicas em função de pressões do ambiente (estresse) seria fundamental. Isto se deve ao fato do ecossistema de pastagem ser extremamente dinâmico, sofrendo constantes interferências, devido a práticas de manejo (por exemplo, controle de plantas daninhas, pastejo, pisoteio e deposição de fezes e urina).
O desempenho de diferentes genótipos é afetado distintamente por fatores como temperatura, umidade, características químicas e físicas do solo etc. Essas respostas afetam a competitividade desses genótipos na pastagem. Assim, o conhecimento da adaptação de genótipos de forrageiras a diferentes condições ambientais é essencial para auxiliar nas decisões de manejo, como a escolha dos cultivares mais apropriados, sistemas de pastejo, adubação etc.
Em pastagens, as práticas de manejo, via de regra, visam a aumentar a eficiência competitiva das plantas forrageiras, diminuindo, assim, a velocidade do processo de sucessão. O objetivo seria buscar e manter composição botânica e cobertura vegetal do solo que garantissem níveis satisfatórios de produtividade animal.
Em áreas pouco alteradas (com baixo nível de intervenção antropogênica), como pastagens nativas utilizadas extensivamente, o clima, principalmente a temperatura e a precipitação pluvial, além das características químicas e físicas do solo são os determinantes principais para a competição. Por outro lado, em pastagens cultivadas, ou em pastagens nativas com alta intensidade de uso, as práticas de manejo (por exemplo, o uso de fertilizantes e herbicidas, a escolha das espécies forrageiras e o regime de pastejo adotado) são os principais fatores que influenciam a competição e, conseqüentemente, as plantas que serão capazes de sobreviver e se desenvolverem sob as condições de crescimento criadas.
As práticas de manejo podem ter grandes impactos na interação competitiva entre espécies e na composição do pasto (Kemp & King, 2001), além de serem capazes de reger a dinâmica do processo de sucessão secundária que se desenvolve na pastagem.
Adubação
A fertilidade do solo desempenha papel importante na capacidade competitiva de espécies vegetais (por exemplo, Suding et al., 2004).
Na pastagem, as espécies diferem quanto as suas capacidades (eficiência) para extrair e utilizar nutrientes, o que resulta in diferenças no crescimento. Na ausência de competição, virtualmente cada espécie vegetal irá mostrar algum aumento na taxa de crescimento com o aumento na fertilidade do solo. A aplicação de fertilizantes irá favorecer de maneira diferenciada aquelas espécies que tiveram o crescimento mais desfavorecido pela baixa fertilidade do solo. Estas espécies seriam aquelas altamente competitivas, com altas taxas máximas de crescimento.
Do ponto de vista competitivo, o objetivo da adubação de pastagens seria aumentar a disponibilidade de um recurso limitante (fertilidade do solo), visando a aumentar a eficiência competitiva das plantas forrageiras e a diminuir a intensidade do processo de sucessão secundária. Por exemplo, uma visão comum é que fertilizantes fosfatados tendem a favorecer as leguminosas, até que o nível de N do solo esteja alto, enquanto que aplicações de N geralmente levam a dominância da gramínea (Kemp & King, 2001). No Brasil, adubações fosfatadas e nitrogenadas teriam importâncias diferenciadas para promover o aumento da competitividade de gramíneas, em pastagens formadas sob áreas originalmente de floresta e de cerrado (Dias-Filho, 2003).
Pastejo
A herbivoria pode alterar a habilidade competitiva das plantas através das mudanças provocadas na morfologia e crescimento e pela influência que exerce na distribuição e abundância das plantas. A herbivoria pode causar ainda inversão na competição, da parte aérea para as raízes, devido à remoção da biomassa aérea, que causa maior entrada de luz no ambiente.
O pastejo pode interferir no padrão de dominância das espécies da pastagem. A área foliar residual após o pastejo seria fator de grande importância para determinar que espécie teria a capacidade de interceptar mais luz e, assim, ter maior chances para rebrotar eficientemente. No entanto, conforme revisado por Nurjaya & Tow (2001), outros fatores, como taxas de aumento da área foliar e os valores de área foliar específica estariam altamente e positivamente correlacionados como a capacidade competitiva.
A “colheita” da forragem que acontece durante o pastejo, dificilmente é feita de forma que otimize a produção da planta forrageira. Isso se deve ao fato do pastejo ser normalmente feito de forma seletiva pelos animais, sendo que as partes da planta preferencialmente consumidas (folhas mais novas, responsáveis por grande parte do desempenho fotossintético da planta) seriam aquelas que teriam maior impacto positivo na capacidade produtiva da planta forrageia. A influência do animal na pastagem não se limita somente ao consumo da forragem. A constante deposição desordenada de esterco e urina e o pisoteio têm a capacidade de afetar o desempenho da pastagem por afetar processos complexos como o ciclo de nutrientes (Dias Filho et al., 2001; Dias Filho, 2003).
A forma de funcionamento e os efeitos da competição e sucessão em pastagens diferem de outros ecossistemas vegetais naturais ou antropogênicas. As principais diferenças são causadas pela presença do animal de pastejo. O pastejo e o pisoteio reduzem a altura e, algumas vezes a densidade do dossel. Isto reduz competição por luz entre ramos e pode indiretamente reduzir a intensidade da competição radicular se parte do sistema radicular reagir à desfolhação crescendo mais lentamente ou morrendo.
A influência do pastejo é crítica para a composição e produtividade da pastagem. Controlar o consumo animal total e a habilidade dos animais selecionarem a sua dieta, através da manipulação da carga animal, é uma das ferramentas mais poderosas que os produtores possuem para manejar a composição da pastagem (Kemp & King, 2001).
A desfolha devido ao pastejo ou corte reduz a vantagem competitiva adquirida pelas plantas que emergiram mais cedo, que têm um embrião maior para iniciar o crescimento e que têm atributos favoráveis como maiores taxas iniciais de crescimento relativo, taxas de perfilhamento, expansão foliar, expansão radicular e estatura (Tow & Lazenby, 2001). Os herbívoros mudam o equilíbrio da competição por consumirem preferencialmente os competidores dominantes, alterando a habilidade competitiva relativa das espécies e promovendo a abundância de plantas que são menos comuns (Peltzer & Wilson, 2001).
Segundo Peltzer & Wilson (2001), o pastejo pode ter maior impacto em habitats menos produtivos em função da menor disponibilidade de nutrientes no solo desses habitats, para serem utilizados pelas plantas para a rebrota e devido ao efeito deletério que o pastejo teria sobre a associação de micorrizas com as raízes das plantas pastejadas, prejudicando a absorção dos nutrientes necessários para a recuperação da planta.
Em pastagens, o impacto de altas pressões de pastejo pode ser suficientemente intenso para provocar danos severos à parte aérea e subterrânea da vegetação, sendo capaz de provocar reversão na sucessão a uma condição de solo nu.
Controle de invasoras
A presença de plantas daninhas na pastagem é sempre reflexo de práticas anteriores, onde o manejo inapropriado levou a que as espécies desejáveis se tornassem menos competitivas (Kemp & King, 2001).
Plantas que se estabelecem precocemente não têm somente grande efeito adverso nas plantas que se estabelecem mais tarde, mas, por sua vez, são pouco afetadas pelas plantas de estabelecimento tardio. Dessa forma as plantas de estabelecimento precoce tendem a persistir. Esse precipício é normalmente usado no manejo das plantas daninhas de pastagem (Tow & Lazenby, 2001). Uma situação típica seria durante a fase de estabelecimento, daí a importância de usar sementes de bom valor cultural e garantir preparo adequado da área.
Quando áreas de solo descoberto são criadas na pastagem, o caminho estaria aberto para o estabelecimento de plantas oportunistas (invasoras de pastagens) (Dias-Filho, 1990; 2003). O controle de plantas daninhas em pastagem tem como objetivo principal manejar a área, visando a incentivar o desenvolvimento da pastagem, suplantando, assim, o das plantas daninhas em dado tempo e local (Dias-Filho, 1990). A lógica para isso é que a proliferação das plantas daninhas na pastagem seria conseqüência da queda de produtividade (degradação) desta pastagem e não a causa desse processo (Dias-Filho, 2003). Desta forma, a manutenção da pastagem mais vigorosa causaria a redução dos espaços físicos (por exemplo, áreas de solo sem a proteção da vegetação) ideais para a germinação das sementes ou para a rebrota de estruturas vegetativas das plantas daninhas, dificultando ainda o desenvolvimento das plantas daninhas já formadas.
O fogo é um dos métodos mais comumente empregados como forma de desacelerar o curso da sucessão secundária em pastagens tropicais. Ao contrário de outros métodos de controle de plantas invasoras de pastagens, o fogo não é seletivo, atingindo indiscriminadamente as plantas “desejáveis” e as “indesejáveis”. A resposta das plantas ao fogo depende de vários fatores, dentre os quais destacam-se a morfologia, a fenologia e a época de aplicação do fogo. A lógica empregada na queima de pastagens é que a gramínea seria tolerante ao fogo, rebrotando imediatamente após, ganhando assim, vantagem competitiva perante as dicotiledôneas que rebrotariam mais lentamente, ou não rebrotariam. Em termos competitivos, em curto prazo, o principal benefício da queima da pastagem seria o aumento na disponibilidade de luz para o desenvolvimento do capim. A competição por outros tipos de recursos, como nutrientes e espaço poderia não ser afetada, já que muitas dicotiledôneas lenhosas (basicamente, aquelas que são capazes de regeneração vegetativa) não seriam afetadas, a longo prazo, pelo fogo, pois rebrotariam após a queima. Para estas plantas, o fogo teria o mesmo efeito da roçagem. Portanto, em pastagens que são freqüentemente queimadas e que, posteriormente, não são submetidas a outros métodos de controle, ou a outras formas de manejo que aumentem a capacidade competitiva das forrageiras (como adubações e controle da pressão de pastejo), o percentual de plantas daninhas pode aumentar, passando a comunidade dessas plantas a ser constituída, quase que exclusivamente, por plantas que apresentam alguma forma de tolerância à queima (Dias Filho, 2003).
Dependendo da sua intensidade e duração, o fogo é capaz de destruir as sementes de plantas daninhas, que ainda estejam presentes nos frutos, presos às plantas, e também, muitas das sementes que se encontram na superfície, ou muito próximas à superfície do solo. No entanto, em alguns casos, a queima pode atuar como agente responsável pela quebra de dormência, estimulando a germinação das sementes de plantas daninhas no banco de semente do solo. Isto se deve ao efeito do calor, no rompimento de tegumentos impermeáveis das sementes, ou em função da ação de substâncias químicas, presentes na fumaça, oriunda da biomassa vegetal queimada (revisado por Gallagher et al. 1999). Por outro lado, as cinzas aumentariam o conteúdo de nitrogênio do solo, elevando, conseqüentemente, os níveis de nitrato, que poderia estimular a germinação de certas sementes dormentes.
Em função do custo relativamente baixo e da facilidade de aplicação, a queima da pastagem, como forma de controle de plantas daninhas, é muitas vezes utilizada exageradamente, podendo causar, a médio e longo prazos, mais danos do que benefícios para a produtividade da pastagem.
Sucessão em pastagens
Todas as comunidades vegetais estão sujeitas a mudanças na composição de espécies e na importância relativa de formas de vida vegetal (ervas, arbustos, árvores) através do tempo (Grime, 2001). Na sucessão existe alteração progressiva na estrutura da vegetação e nas espécies dominantes. Na sucessão primária, um novo hábitat, normalmente carecendo de solo e vegetação, é colonizado (como em algumas áreas de mineração). Na sucessão secundária, existe a recolonização de um hábitat alterado (como em pastagens cultivadas).
Em pastagens nativas, os padrões de sucessão e competição geralmente são influenciados pelo impacto causado pelo pastejo, pisoteio e deposição de fezes e urina (Skarpe, 2001). A presença do herbívoro provoca alterações, por exemplo, na estrutura da vegetação, devido ao pastejo seletivo e ao pisoteio e na ciclagem de nutrientes, em função da deposição irregular de fezes e urina. O pastejo pode ainda ter impacto indireto na intensidade do fogo, na ocasião de eventuais queimas da pastagem, afetando o curso da sucessão. A razão para isso seria que o pastejo provocaria redução na biomassa de gramíneas, o principal combustível para a propagação do fogo em pastagens nativas. Com a diminuição na intensidade do fogo, espécies menos tolerantes a queima poderiam ser beneficiadas, alterando a composição da vegetação (revisado por Skarpe, 2001).
Em pastagens cultivadas, o processo natural de sucessão secundária sofre constantes intervenções de origem antropogênica, que têm o objetivo de desacelerar a sucessão através de práticas de manejo que visam a controlar plantas invasoras e a manter a pastagem apenas com as espécies forrageiras (normalmente exótica àquele ambiente) de interesse direto para a alimentação animal. Dependendo da vegetação original (floresta, cerrado etc.) e das técnicas empregadas no preparo da área e na implantação da pastagem, o processo de sucessão secundária, que tem início imediatamente após o preparo da área (após a retirada da vegetação original), poderá ter diferentes intensidades. Portanto, o esforço (tempo, energia etc.) gasto pelo produtor para manter a pastagem livre de plantas indesejáveis (invasoras, daninhas), desde a sua implantação, será função da força do processo de sucessão. A força do processo de sucessão secundária (recolonização da pastagem) por sua vez dependerá, principalmente, do tamanho do banco de sementes e propágulos existente no solo, da “taxa de reposição” desse banco, através da migração ou dispersão de propágulos e de novas sementes via dispersores e da eficiência competitiva da(s) planta(s) forrageira(s) (geralmente uma única espécie). A “eficiência competitiva” da forrageira será, em grande parte, resultante de decisões de manejo, as quais incluem dentre outras, a escolha da espécie forrageira mais adaptada, adubação, duração e pressão do pastejo etc.
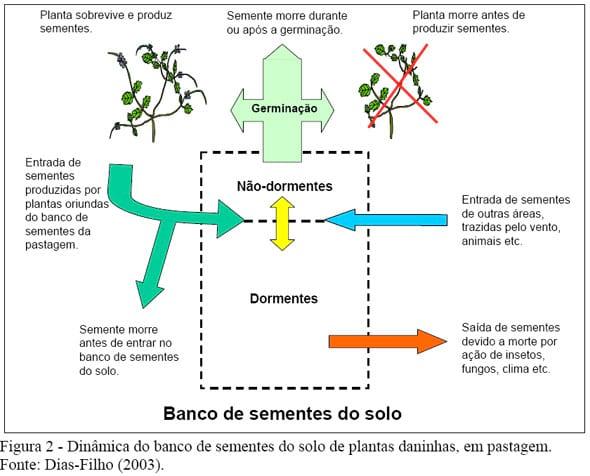
O banco de sementes de pastagens cultivadas tem dinâmica muito peculiar (Figura 2). De acordo com Dias Filho (2003), em pastagens, o tamanho e composição do banco de sementes do solo estão normalmente relacionados com o manejo e a produtividade da pastagem. Por exemplo, quanto maior for, ou tiver sido o período de infestação da pastagem por plantas daninhas, maior e mais diversificado deverá ser o seu banco de sementes. Por outro lado, conforme representado na Figura 2, sementes de plantas daninhas de locais distantes podem também migrar de outros locais para o banco de sementes do solo de determinada pastagem, trazidas pelo vento, animais (pássaros, morcegos etc.), máquinas agrícolas, entre outros, através de fenômeno chamado “dispersão de sementes”. Nesse caso, o manejo da pastagem teria influência limitada para controlar a entrada de sementes. O gado pode também introduzir (dispersar) sementes de plantas daninhas na pastagem, de forma ativa, através das fezes (Jones et al. 1991), que conteriam sementes das plantas consumidas em outras locais, ou mesmo, passivamente, transportando sementes aderidas ao pêlo ou às patas. Estudos mostram que as sementes ingeridas pelo gado começam a ser excretadas 24 horas após a ingestão, continuando a aparecer nas fezes até cinco dias após (Stanton et al. 2002). Sementes de plantas daninhas também podem entrar na área, durante a fase de estabelecimento da pastagem, como contaminadoras de lotes de sementes de capim, com baixo grau de pureza (baixo valor cultural), ou aderidas a máquinas agrícolas e as roupas e calçados das pessoas.
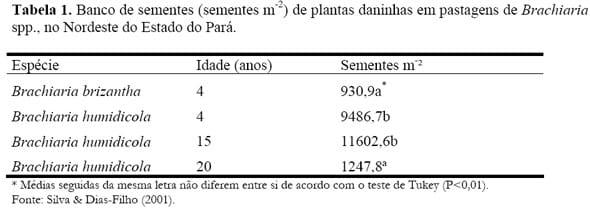
Levantamento feito no banco de sementes de pastagens de Brachiaria spp., no nordeste do Estado do Pará, encontrou-se bancos de sementes de plantas daninhas com tamanhos que variaram de em torno de 1.000, a até mais de 10.000 sementes por metro quadrado de solo (até 10 cm de profundidade) (Tabela 1) (Silva & Dias-Filho, 2001).
No solo, as sementes vivas das plantas daninhas podem encontrar-se em dois estados distintos: dormentes e não-dormentes. As sementes não-dormentes podem germinar imediatamente, quando encontram condições ambientais (luz, umidade, temperatura etc.) adequadas, ou entrar em estado de dormência, quando as condições são inadequadas para a germinação.
Algumas sementes, no entanto, já são produzidas dormentes, não germinando imediatamente, mesmo sob condições ambientais favoráveis. As sementes dormentes são as que formam o banco de sementes do solo, sendo que as sementes não-dormentes estão presentes no banco, sob forma transitória (Figura 2).
Segundo Dias Filho (2003), em pastagens, as duas principais causas da dormência das sementes são de natureza física e fisiológica. Na dormência provocada por razões físicas, o tegumento que envolve a semente impede a embebição (troca de água) ou a troca de gases pela semente com o meio-ambiente, ou a expansão do embrião, no interior da semente.
Sementes que apresentam esse tipo de dormência são chamadas sementes duras. Na dormência fisiológica, inibidores da germinação, encontrados no tegumento da semente ou no próprio embrião, impedem a germinação, por bloquearem processos metabólicos internos da semente.
O tempo que a semente permanece viável no solo, em estado de dormência, pode variar bastante dentre espécies de plantas daninhas e tipos de ecossistemas. Quanto maior o tempo e o percentual de sobrevivência das sementes no solo, maior será o potencial da planta daninha em contribuir para aumentar o banco de sementes do solo.
O processo natural de quebra da dormência das sementes no solo é influenciado por uma série de fatores, inerentes às causas da dormência, espécie vegetal, ou características ambientais, sendo que o tempo necessário para esse processo pode variar de algumas horas a até vários anos. Dentre os fatores responsáveis pela quebra de dormência, estariam substâncias químicas encontradas no solo; as variações bruscas na temperatura e no teor de umidade do solo (como as causadas durante a queima da pastagem, ou pela deficiência na cobertura vegetal do solo); distúrbios sofridos pelo solo (como os provocados pela gradagem do solo), que causariam a movimentação vertical das sementes no perfil do solo, expondo, conseqüentemente, muitas dessas sementes a condições ambientais mais favoráveis para a quebra de dormência, como a variações dos níveis diários de temperatura, luz e umidade do solo.
Algumas substâncias químicas, presentes no solo, podem estimular (ou inibir) a germinação das sementes e, assim, contribuir para a quebra (ou manutenção) da dormência. Duas dessas substâncias, que têm sido muito estudadas, na germinação de espécies tropicais e subtropicais, são o etileno e o nitrato (revisado por Carmona, 1992; Karssen & Hilhorst, 1992). O etileno é produzido por microrganismos do solo e pelas raízes de algumas plantas; podendo estimular ou inibir a germinação das sementes no solo. O nitrato ocorre naturalmente no solo e faz parte do ciclo do nitrogênio. Assim, a decomposição da matéria orgânica do solo ou qualquer evento que altere a população de macro e microrganismos nesse solo, teria influência direta nos níveis de nitrato. Os mecanismos bioquímicos responsáveis pela ação do nitrato, na quebra de dormência, são complexos e ainda pouco entendidos (Karssen & Hilhorst, 1992). Do ponto de vista ecológico, a ação do nitrato, em sementes não expostas à luz, tem sido proposta como indicador natural, para a semente, da diminuição da cobertura vegetal do solo (Pons, 1989). A lógica seria que, maiores níveis de nitrato, no solo, significariam menor quantidade de plantas (mais plantas = maior absorção do nitrato do solo) e, portanto, menor competição para a plântula recém-formada.
Uso da sucessão vegetal na recuperação de pastagens degradadas
Dias Filho (2003) propõe o uso da sucessão secundária, através do pousio da pastagem, como forma alternativa de recuperação de áreas de pastagem degradada. Essa alternativa seria indicada principalmente recuperar áreas que não deveriam ter sido originalmente desmatadas, como aquelas localizadas ao longo de cursos d’água, ou sobre solos considerados inapropriados para atividades agropecuárias. O processo consistiria simplesmente no abandono da área, por tempo indefinido, podendo ou não ser reutilizada, no futuro, para nova formação (renovação) da pastagem, ou para outro fim agropecuário ou florestal. No caso de pastagens formadas em áreas originalmente sob floresta, após o abandono, o local passa a ser paulatinamente invadido por arbustos e árvores, que, com o decorrer do tempo, podem levar a formação de vegetação característica de floresta secundária. Nestas florestas secundárias, muitas funções da floresta primária podem ser parcialmente restabelecidas. As principais seriam: a intensificação do ciclo hidrológico, devido à capacidade da floresta secundária em absorver água de camadas mais profundas do solo; a atuação como sumidouro do carbono atmosférico, através da fixação do gás carbônico do ar, durante a fotossíntese e a conservação de nutrientes, através da transferência de nutrientes do solo (onde estariam mais vulneráveis à erosão e à lixiviação) para a biomassa da floresta secundária (Guggenberger & Zech, 1999; Nepstad et al. 2001).
Segundo Nepstad et al. (2001), a taxa média de acúmulo de biomassa aérea, em florestas secundárias, na Amazônia, seria de quatro toneladas por hectare por ano. Em geral, o processo de sucessão vegetal, em pastagens abandonadas, seria mais lento do que o que ocorreria em áreas desmatadas e imediatamente abandonadas (Mesquita et al. 2001); ou mesmo em áreas originalmente sob agricultura migratória (Moran et al. 2000). Os fatores que influenciam a taxa de formação da floresta secundária, em pastagem abandonada, ainda são pouco entendidos. No entanto, a baixa disponibilidade no solo de sementes e propágulos, a alta taxa de predação de sementes e plântulas, a baixa fertilidade do solo, a competição com o capim remanescente e o microclima têm sido propostas como as principais barreiras para a formação da floresta secundária, em pastagens abandonadas (Holl, 1998; 2002; Nepstad et al.1991; 1996; Slocum, 2001; Uhl et al. 1988; Wijdeven & Kuzee, 2000). Sabe-se, no entanto, que a taxa de ocupação da vegetação secundária nessas áreas, estaria inversamente relacionada com a intensidade de uso da pastagem original (Uhl et al. 1988; Nepstad et al. 1996). Portanto, pastagens que sofreram formas mais agressivas de uso, como aquelas causadas pelo emprego de queimas freqüentes, superpastejo, aplicação sistemática de herbicidas ou uso freqüente de mecanização, para o controle de plantas daninhas, quando abandonadas, apresentariam taxa de ocupação pela vegetação secundária mais lenta do que as pastagens que foram utilizadas com menor intensidade. A razão disto é que nas pastagens que sofreram utilização agressiva, o banco de sementes do solo não seria tão grande e diversificado, como nas áreas utilizadas menos intensamente. Por outro lado, nestes locais, outras formas de regeneração da vegetação secundária, como a rebrota de raízes, bulbos ou de outras estruturas vegetativas, teriam sofrido maior taxa de esgotamento, devido à ação mais intensa de formas de controle, como a aplicação de herbicidas, queima e roçagens freqüentes. Finalmente, em muitas destas áreas, os níveis de fertilidade do solo seriam relativamente baixos e a compactação do solo seria grande. Essas características atrasariam a taxa de ocupação e a velocidade de desenvolvimento da vegetação secundária.
De acordo com Dias Filho (2003), alternativamente, o processo natural de sucessão da vegetação secundária, em pastagens degradadas abandonadas, poderia sofrer intervenções, através do controle seletivo (por exemplo, através do raleamento) da vegetação secundária natural, visando a dificultar o desenvolvimento de plantas indesejáveis e facilitar o estabelecimento das plantas consideradas desejáveis.
Outra forma de manejo da vegetação secundária, para a recuperação de áreas degradadas, seria o plantio estratégico de espécies, com superior capacidade de crescimento e acúmulo de biomassa e de nutrientes, ou de maior valor econômico, em sistemas conhecidos, respectivamente, por “melhoramento” ou “enriquecimento” da vegetação secundária (Sanchez, 1999). Esses sistemas de manejo vêm sendo recomendados e utilizados, em várias regiões tropicais e subtropicais (por exemplo, Brienza Júnior et al. 1998; Gallagher et al. 1999; Otsamo, 2000; Roder & Maniphone, 1998; Szott et al. 1999), sendo ainda indicados como formas de superar as barreiras naturais para a regeneração da floresta, em pastagens abandonadas (Zanne & Chapman, 2001; Zimmerman et al. 2000), ou para restabelecer composição da floresta primária, em florestas secundárias (Aide et al. 2000).
No caso de pastagens degradadas abandonadas, os objetivos do melhoramento e do enriquecimento da vegetação secundária são:
1. aumentar a velocidade e a eficiência do processo de sucessão, através do plantio de espécies de crescimento rápido, com alta capacidade de acúmulo de biomassa, capazes de atrair animais dispersores de sementes e de propiciar condições ambientais que possam facilitar o desenvolvimento das demais espécies vegetais, no processo de sucessão da floresta (Ganade & Brown, 2002; Holl, 2002; Slocum, 2001; Zanne & Chaman, 2001); e
2. agregar valor econômico à área, através do plantio de espécies com potencial para exploração agrícola, como espécies frutíferas ou medicinais, ou para a exploração de madeira, como espécies florestais.
Tópicos relacionados
Autores:

Junte-se para comentar.
Uma vez que se junte ao Engormix, você poderá participar de todos os conteúdos e fóruns.
* Dados obrigatórios
Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.
Criar uma publicaçãoVocê pode estar interessado em
