Uso de aditivo alimentar equilibrador da flora intestinal em aves de corte e de postura
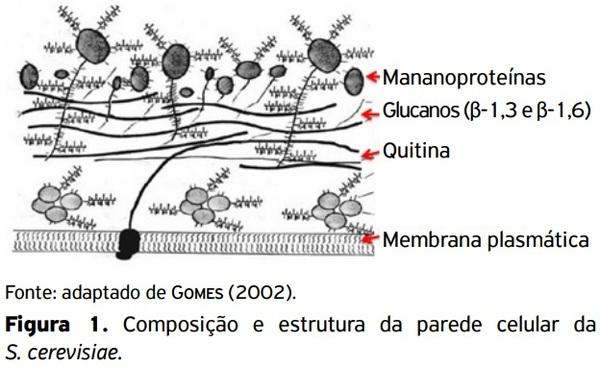
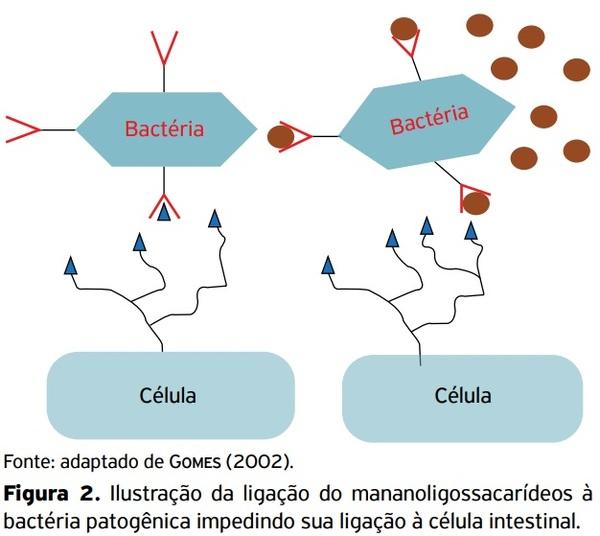
ALLES, M.S.; HARTEMINK, R.; MEYBOOM, S.; HARRYVAN, J.L.;
VAN LAERE, K.M.; NAGENGAST, F.M.; HAUTVAST, J.G. Effect of transgalactooligosaccharides on the composition of the human intestinal microflora and on putative risk markers for colon cancer.
The American Journal of Clinical Nutrition, v.69, p.980-991, 1999.
AYANWALE, B.A.; KPE, M.; AYANWALE, V.A. The effect of supplementing saccharomyces cerevisiae in the diets on egg laying and egg quality characteristics of pullets. International
Journal of Poultry Science, v.5, p.759-763, 2006.
BIGGS, P.; PARSONS, C.M.; FAHEY, G.C. Effects of several oligosaccharides on growth performance, nutrient digestibilities, and caecal microbial populations in young chicks. Poultry Science, v.86, p.2327-2336, 2007.
BOUHNIK, Y.; FLOURIE, B.; D’AGAY-ABENSOUR, L.; POCHART,
P.; GRAMET, G.; DURAND, M.; RAMBAUD, J.C. Administration of transgalactooligosaccharides increases fecal bifidobacteria and modifies colonic fermentation metabolism in healthy humans.
Journal of Nutrition, v.127, p.444-448, 1997.
CASTANON, J.I.R. History of the use of antibiotic as growth promoters in European Poultry Feeds. Poultry Science, v.86, p.2466-2471, 2007.
COMPÊNDIO BRASILEIRO DE ALIMENTAÇÃO ANIMAL. Sumário:
Guia de aditivos. São Paulo: Sindicações, 2009.
COSTA, F.G.P.; NOBRE, I.S.; SILVA, L.P.G.; GOULART, C.C.;
FIGUEIREDO, D.F.; RODRIGUES, V.P. The use of prebiotic and organic minerals in rations for japonese laying quail. International
Journal of Poultry Science, v.7, n.4, p.339-343, 2008.
COTTER, P.F. Modulation of immune response: current perceptions and future prospects with an example from poultry.
In: ALLTECH’S ANNUAL SYMPOSIUM ON 90 BIOTECHNOLOGY
IN FEED INDUSTRY, 10., 1994, Nottingham. Proceedings...
Loughborough, UK: Nottingham University Press, 1994. p.105-203.
DIMOVELIS, P.; CHRISTAKE, E.; TSEVERINI-GOUSSI, A.; SPAIS,
A.B. Effect of Bio-Mos on growth, egg production and egg quality of Lohmann brown layers. In: ALLTECH’S ANNUAL NUTRITIONAL
BIOTECHNOLOGY IN THE FEED AND FOOD INDUSTRIES, 19.,
2003, Lexiton. Proceedings... Lexiton, Ky: [s.n.], 2003.
DIONÍZIO, M.A.; BERTECHINI, A.G.; KATO, R.K.; TEIXEIRA, A.S.
Prebiótico como promotor de crescimento para frangos de corte – desempenho e rendimento de carcaça. Ciência Agrotécnica, n.3, p.1580-1587, 2002.
DONALSON, L.M.; KIM, W.K.; CHALOVA, V.I.; HERRERA, P.;
WOODWARD, C.L.; MCREYNOLDS, J.L.; KUBENA, L.F.; NISBET,
D.J.; RICKE, S.C. In vitro anaerobic incubation of Salmonella enterica serotype Typhimurium and laying hen cecal bacteria in poultry feed substrates and a fructooligosaccharide prebiotic.
Anaerobe, Iowa, v.13, n.5-6, p.208-214, 2007.
FERREIRA. A.P.; ASTOLFI-FERREIRA, C.S. Medidas inespecíficas para o controle bacteriano. In: SIMPÓSIO BRASIL SUL DE
AVICULTURA, Chapecó, 2006, Anais... Chapecó, 2006. p.56-66.
GARCIA, N.L.; CACHALDORA, P.; TUCKER, L. BAUCELLS, F.; NEDEL,
P. Effect of mannanoligossacharides suplementation to laying hen diets. Poultry Science, n.83, Supplement 1, p.397, 2004.
GIBSON, G.R.; ROBERFROID M.B. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. Journal of Nutrition, v.125, n.6, p.1401-1412, 1995.
GOMES, M.A.B. Aditivos Probióticos, Prebióticos e Simbióticos na Alimentação Animal. Disponível em: <www.mariboi.com.br/_ assets/artigos/13_artigo.pdf, 2002>. Acesso em: 15 abr. 2011.
GOSH, H.K.; HALDER, G.; SAMANTA, G.; PAUL, S.K.; PYNE,
S.K. Effect of dietary supplementation of organic acid and mannanoligosaccharide on the performance and gut health of japonese quail (Coturnix coturnix japonica). Asian Journal of
Poultry Science, v.1, n.1, p.1-7, 2007.
HOOGE, D. Dietary alternatives for improving live performance of antibiotic-free poultry. International Journal of Poultry Science, v.3, p.163-174, 2004.
HOSSEINI, S.A.; LOTFOLLAHIAN, H.; KAMYABAND, A.; MADHAVI, A
Study on the effect of yeast (Saccharomyces cerevisiae) utilization on the commercial layer hen’s performance. Pakistan Journal of
Biological Sciences, v.12, p.2346-2349, 2006.
IBRAHIM, Z.A. Modulation of immunity and some biological functions of japonese quail by mannan oligosaccharide and B-glucan administration. Egypt Poultry Science, v.31, p.867-882, 2011.
IJI, P.A.; TIVEY, D.R. Natural and synthetic oligosaccharides in broiler chicken diets. World Poultry Science Journal, v.54, p.129-143, 1998.
IJI, P.A.; SAKI, A.A.; TIVEY, D.R. Intestinal structure and function of broiler chickens on diets supplemented with a mannanoligosaccharide. Journal of the Science of Food and
Agriculture, v.81, n.12, p.1181-1192, 2001.
JUNG, S.J.; HOUDE, R.; BAURHOO, B.; ZHAO, X.; LEE, B.H. Effects of galacto-oligosaccharides and a Bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens. Poultry science, v.87, n.9, p.1694-1699, 2008.
KNUDSEN, K.E.B.; HESSOV, I. Recovery of inulin from Jerusalem artichoke (Helianthus tuberosus L.) in the small intestine of man.
British Journal of Nutrition, v.74, p.101-113, 1995.
KOGAN, G.; KOCHER, A. Role of yeast cell wall polysaccharides in pig nutrition and health protection. Livestock Science, v.109, p.161-165, 2007.
KRUGER, M.C.; BROWN, K.E.; COLLETT, G.; LAYTON, L.; SCHOLLUM,
L.M. The effect of fructooligosaccharides with various degrees of polymerization on calcium bioavailability in the growing rat.
Experimental Biology and Medicine, v.228, p.683-688, 2003.
LEMOS, M.J.; CALIXTO, L.F.L.; NASCIMENTO, A.A.; SALES,
A.; SANTOS, M.A.; AROUCHA, R.J.N. Morfologia do epitélio intestinal de codornas japonesas alimentadas com parede celular da Saccharomyces cerevisiae. Ciência Rural, v.43, n.12, p.2221-2227, 2013.
LEMOS, M.J.; CALIXTO, L.F.L.; LIMA, C.A.R.; REIS, T.L.; REGO,
R.S.; NAK, S.Y.; AROUCHA, R.J.N. Níveis de prebiótico na dieta sobre o desempenho e a qualidade de ovos de codornas japonesas. Revista Brasileira de Saúde e Produção Animal, v.15, n.3, p.614-626, 2014.
LODDI, M.M. Probióticos, prebióticos e acidificantes orgânicos em dietas para frangos de corte. Tese (Doutorado em Zootecnia) –
Universidade Estadual Paulista “Júlio de Mesquita Filho”,
Jaboticabal, 2003.
MACARI, M.; MAIORKA, A. Função gastrintestinal e seu impacto no rendimento avícola. In: CONFERÊNCIA APINCO DE CIÊNCIA E
TECNOLOGIA AVÍCOLAS, 2., 2000, Campinas. Anais... Campinas:
FACTA, 2000. p.161-174.
MACARI, M.; FURLAN, R.L. Probióticos. In: CONFERÊNCIA APINCO
DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1., 2005, Campinas.
Anais... Campinas: FACTA, 2005. p.53-71.
MAIORKA, A.; SANTIN, E.; SUGETA, S.M.; ALMEIDA, J.G.; MACARI, M.
Utilização de prebióticos, probióticos ou simbióticos em dietas para frangos. Revista Brasileira de Ciência Avícola, v.3, p.75-82, 2001.
MARKOVIC, R.; SEFER, D.; KRSTIC, M.; PETRUJKIC, B. Effect of different growth promoters on broiler performance and gut morphology.
Archivos de Medicina Veterinária, v.41, p.163-169, 2009.
MATHEUS, K.M.; BERNSTEIN, J.R.; BUZBY, J.C. International Trade of Meat/Poultry Products and Food Safety Issues. Washington, DC:
Economic Research Service/USDA, 2003. p.49-73.
MORALES-LÓPEZ, R.; AUCLAIR, E.; GARCÍA, F.; ESTEVE-GARCIA,
E.; BRUFAU,
J. Use of yeast cell walls; β-1,3/β-1,6 glucans; and mannoproteins in broiler chicken diets. Poultry Science, v.88, p.601-607, 2009.
NEWMAN, K. Form follows function in picking MOS product.
Feedstuffs, v.79, n.4, 2007.
NUMAZAKI, E.M. Adição de mananoligossacarídeos e halquinol em dieta de poedeiras Bovans White. Dissertação (Mestre em Ciências
Agrárias) – Universidade de Brasília, 2008.
NUNES, A.D. Influência do uso de aditivos alternativos a antimicrobianos sobre o desempenho, morfologia intestinal e imunidade de frangos de corte. Dissertação (Mestrado em Medicina Veterinária) – Universidade de São Paulo,
São Paulo, 2008.
NUNES, A.D.; VAZ, A.C.N.; RASPANTINI, L.E.; SILVA, E.M.;
ALBUQUERQUE, R. Desempenho e morfologia intestinal de frangos de corte alimentados com rações contendo aditivos alternativos a antimicrobianos. Brazilian Journal of Veterinary
Research and Animal Science, v.46, n.6, 2009.
OLIVEIRA, M.C.; MACHADO, M.G.; GONÇALVES, B.N.; MACEDO,
C.M.R.; ASSIS, F.A. Dietas com mananoligossacarídeo e níveis reduzidos de cálcio para codornas japonesas. Revista Brasileira de Zootecnia, v.38, n.11, p.2193-2197, 2009.
OTUTUMI, L.K.; FURLAN, A.C.; NATALI, M.R.M.; MARTINS, E.N.M.;
LODDI, M.M.; OLIVEIRA, A.F.G. Utilização de probiótico em rações com diferentes níveis de proteína sobre o comprimento e a morfometria do intestino delgado de codornas de corte. Acta
Scientiarum. Animal Sciences, v.30, n.3, p.283-289, 2008.
PELICANO, E.R.L.; SOUZA, P.A.; SOUZA, H.B.A. Prebióticos e probióticos na nutrição de aves. Revista Ciências Agrárias e da
Saúde, v.2, n.1 p.59-64, 2002.
PELÍCIA, K. Efeito de promotores biológicos e químicos sobre o desempenho, rendimento de carcaça e qualidade da carne em frangos de corte tipo colonial. Dissertação (Mestrado em Zootecnia) –
Universidade Estadual Paulista, Botucatu, 2004.
RIBEIRO, C.L.G.; RUTZ, F.; DALLMANN, P.R.; ZAUK, N.F.; SILVEIRA,
M.H.D.; GONÇALVES, R.A.S.; ANCIUTI, M.A.; ROSSI, P. Efeito da utilização de mananoligossacarídeos (MOS) e de ácidos orgânicos associados à MOS, com e sem antibióticos, na dieta de poedeiras produtoras de ovos avermelhados. Ciência Animal Brasileira, v.11, n.2, p.292-300, 2010.
ROWLAND, I.R. Metabolic interactions in the gut. In: FULLER, R. (Ed.). Probiotics: the scientific basis. London: Chapman and Hall,
1992. p.29-53.
SALYERS, A.A. Agricultural use of antibiotics and antibioticresistance in human pathogens: is there a link? In: ALLTECH.SANNUAL
SYMPOSIUM, 15., Nottingham. Proceedings... Nottingham: Alltech,
1999. p.155-171.
SHANE, M.S. Mannan oligosaccharides in poultry nutrition: mechanism and benefits. Science and technology in the feed industry. Nottingham, UK: Nottingham, 2001. p.65-77.
SANTIN, E.; MAIORKA, A.; MACARI, M.; GRECCO, M.; SANCHEZ,
J.C.; OKADA, T. M.; MYASAKA, A.M. Performance and intestinal mucosa development of broiler chickens fed diets containing
Saccharomyces cerevisiae cell wall. Journal Applied Poultry
Research, v.10, p.236-244, 2001.
SAVAGE, T.F.; ZAKREWSKA, E.I.; ANDREASEN, J.R. The effects of feeding mannan oligosaccharide supplemented diets to poultrys on performance and the morphology of the small intestine. Poultry
Science, v.76, p.139, 1997.
SCAPINELLO, C.; FARIA, H.G.; FURLAN, A.L.; MICHELAN, A.C. Efeito da utilização de oligossacarídeo manose e acidificantes sobre o desempenho de coelhos em crescimento. Revista Brasileira de
Zootecnia, v.30, p.1272-1277, 2001.
SILVA, L.P.; NORBERG, J.L. Prebióticos na nutrição de não ruminantes. Ciência Rural, v.33, n.5, p.983-990, 2003.
SPRING, P.; WENK, C.; DAWSON, A.; NEWMAN, K.E. The effect of dietary mannanoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of Salmonellachallenged broiler chicks. Poultry Science, Champaign, v.79, n.2, p.205-211, 2000.
SPRING, P.; PIRVULESCU, M. Mannanoligosaccharide: Its logical role as natural feed additive for piglets. In: BIOTECHNOLOGY IN
THE FEED INDUSTRY ANNUAL SYMPOSIUM, 13, Norttingham.
Proceedings... Norttingham: Norttingham University Press, p.553, 1998.
SUZUKI, T.; HARA, H. Various non-digestible saccharides increase intracellular calcium ion concentration in rat small-intestinal enterocytes. British Journal Nutrition, v.92, p.751-755, 2004.
YANG, Y.; IJI, P.A.; KOCHER, A.; MIKKELSEN, L.L.; CHOCT, M.
Effects of mannanoligosaccharide on growth performance, the development of gut microflora, and gut function of broiler chickens raised on new litter. Journal Applied Poultry Research, v.16, p.280-288, 2007.
YOUSEFI, M.; KARKOODI, K. Effect of probiotic ThepaxR and
Saccharomyces cerevisiae supplementation on performance and egg quality of laying hens. International Journal of Poultry Science, v.6, n.1, p.52-54, 2007.
ZAFAR, T.A.; WEAVER, C.M.; ZHAO, Y.; MARTIN, B.R.; WASTNEY,
M.E. Non digestible oligosaccharides increase calcium absorption and suppress bone resorption in ovariectomized rats. Journal of
Nutrition, v.134, p.399-402, 2004.
ZAGHINI, A.; MARTELLI, G.; RONCADA, P.; SIMIOLI, M.; RIZZI,
L. Mannanoligosaccharides and Aflatoxin B1 in feed forlaying hens: effects on egg quality, Aflatoxins B1 and M1 residues in eggs, and Aflatoxin B1 levels in liver. Poultry Science, v.84, p.825-832, 2005.
XU, Z.R.; HU, C.H.; XIA, M.S.; ZHAN, X.A.; WANG, M.Q. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poultry
Science, v.82, p.1030-1036, 2003.







.jpg&w=3840&q=75)