Nutrição de aves
Novos avanços da nutrição de aves
Publicado: 2 de maio de 2012
Por: Fernando Guilherme Perazzo Costa*, José Humberto Vilar da Silva, Denise Fontana Figueiredo-Lima, Rodrigo Barbosa Lima, Cláudia de Castro Goulart
INTRODUÇÃO
Todas as projeções para o crescimento na produção de carnes apontam o Brasil entre os dois maiores produtores mundiais. Segundo as projeções da FAO em curto prazo será necessário um aumento de 19% na produção de carnes em relação à produção de 2006 para atender a demanda de consumo mundial, principalmente de carne de aves (23,1%) e em menor quantidade de carne suína (16,7%). Atualmente, o Brasil ocupa o 2º e 3º lugares no ranking de maiores produtores de carne de frango e suína, respectivamente (Grolli, 2007) correspondendo aos desafios crescentes do mercado consumidor ditados pelos padrões internacionais de segurança alimentar com responsabilidade ambiental.
Os recentes aumentos nos custos da cadeia produtiva de aves e suínos em decorrência da alta nos preços dos ingredientes utilizados na alimentação animal demandam a adoção de estratégias nutricionais que resultem em melhor aproveitamento dos nutrientes com conseqüente aumentando da eficiência produtiva. Assim, diversas idéias inovadoras têm surgido, sendo que a maioria dos trabalhos de pesquisa sobre alimentação e nutrição animal, em geral, estão focados no uso de aditivos bem como no uso de alimentos alternativos, e na promoção destes ingredientes a fim de reduzir custos nestes segmentos.
Aminoácidos cristalinos
As aves sintetizam proteínas que contém 20 L-aminoácidos e utilizam aminoácidos livres para realizar uma grande variedade de funções; no entanto, são incapazes de sintetizar nove desses aminoácidos decido à ausência de enzimas específicas. Esses aminoácidos essenciais, também chamados de indispensáveis, são: arginina, isoleucina, leucina, lisina, metionina, fenilalanina, treonina, triptofano e valina. Histidina, glicina e prolina podem ser sintetizadas pela ave, mas a uma taxa insuficiente para atender às necessidades metabólicas que algumas situações demandam, além de serem considerados essenciais para aves em crescimento. Tirosina e cisteína podem ser sintetizados quando os aminoácidos essenciais precursores fenilalanina e metionina estão em níveis adequados.
Uma proteína que possui um balanço de aminoácidos essenciais que atendem exatamente às exigências das aves, juntamente com nitrogênio suficiente, proveniente de aminoácidos não-essenciais, para permitir a síntese de todos os aminoácidos não-essenciais, é denominada de proteína ideal.
Atualmente, o uso de aminoácidos sintéticos tem como premissa a aplicabilidade do conceito de proteína ideal, visando atender aos requerimentos de aminoácidos para a mantença e produção, sem deficiências ou excessos. Este interesse explica-se pela busca da redução dos custos mediante a diminuição dos níveis de proteína bruta, bem como na redução da quantidade de N excretado, calculada em 8% para cada ponto percentual reduzido no conteúdo de PB (Kerr & Easter 1995). O conceito de proteína ideal foi primeiramente descrito por Mitchell (1964) que tentou produzir uma dieta que atendesse às necessidades das galinhas usando ingredientes purificados. A simulação do perfil de aminoácidos de tais proteínas "ideais" como a albumina e caseína obtiveram, no entanto, apenas sucesso parcial na tentativa de otimizar o crescimento e eficiência alimentar. Da mesma forma, a formulação subseqüente de aminoácidos ligados à composição corporal da ave também falhou em otimizar o crescimento. Foi apenas após modelar a necessidade de aminoácidos para mantença, crescimento e produção de penas que as formulações resultaram em um crescimento ótimo (Leeson & Summers, 2001).
Os resultados da comparação entre uma ração para frangos de corte originalmente formulada em 1997 (Tabela 1; Ajinomoto, 2007) sem a utilização dos aminoácidos cristalinos lisina e treonina e uma ração reformulada utilizando os mesmos ingredientes e níveis nutricionais, contudo adicionando-se L-lisina HCl e Ltreonina mostraram que a adição dos aminoácidos cristalinos reduziu o custo da ração em 5%, e pôde atender às exigências desses aminoácidos, sem deficiências ou excessos, quando comparada à formulação original. A redução da proteína bruta em 12% quando comparamos as duas rações (20,87 vs. 18,30%), obtida a partir da redução da inclusão do farelo de soja em 31% (24,46% vs. 16,83%) e do aumento em 14% da inclusão do milho (65,47% vs. 74,32%), além do benefício econômico, trariam benefícios zootécnicos e ambientais, pelo melhor balanceamento de aminoácidos, maximização da retenção de nitrogênio e de energia e redução das perdas de nutrientes não digeridos e não aproveitados pelo metabolismo.
Tabela 1. Impacto da utilização de aminoácidos industriais sobre a formulação de uma ração comercial de 1997 para frangos de corte machos dos 22 aos 33 dias de idade
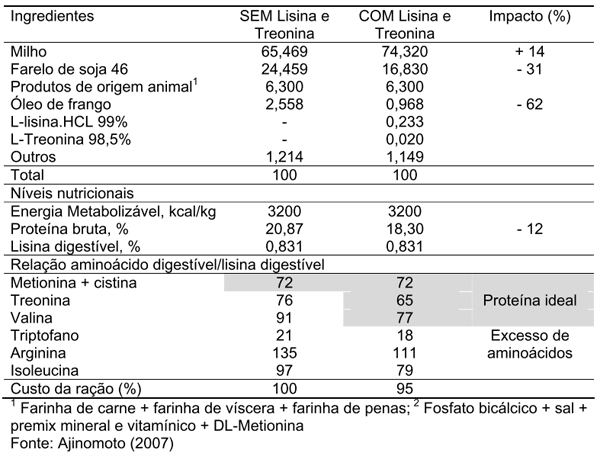
Aftab et al. (2006) afirmam que com a suplementação de aminoácidos cristalinos a PB da dieta poderia ser reduzida em 10% dos níveis recomendados pelo NRC (1994) para cada fase de criação das aves. Por outro lado, Dean et al. (2006) reportaram que o desempenho dos frangos não foi comprometido quando as dietas foram formuladas para conter 75 e 80% da PB e suplementadas com aminoácidos essenciais para atender concentrações similares destes aminoácidos às da dieta controle.
Muitas pesquisas demonstram a possibilidade da redução dos níveis de PB com a adição de aminoácidos sintéticos (Kidd et al., 2000; Costa et al., 2001; Aftab et al., 2006), porém, níveis de redução acima de três pontos percentuais, mesmo com a suplementação dos aminoácidos essenciais metionina, lisina, treonina e triptofano resultaram em pior desempenho de frangos de corte na fase de crescimento (Sabino et al., 2004; Dean et al., 2006). Assim, é razoável considerar que a progressiva redução da proteína bruta dietética pode levar a deficiência nos níveis de treonina e principalmente da valina e isoleucina, geralmente supridos por dietas com alta PB (Peganova & Eder, 2002). Portanto, para o máximo desempenho de aves submetidas a dietas com maiores reduções protéicas, os valores mínimos para isoleucina, valina, arginina e triptofano devem ser estabelecidos quanto às respectivas suplementações consideradas.
Vitaminas
Assim como para a maioria dos nutrientes, as exigências vitamínicas para aves tem provavelmente mostrado pequenas mudanças nos últimos anos (Leeson, 2006). Devido às condições modernas de manejo, as aves são submetidas a diversos fatores estressantes que podem aumentar suas exigências em vitaminas, como detalhado pelo NRC (1994). Logo, tem sido considerada a elevação dos níveis de inclusão dietéticas das vitaminas de modo a fornecer uma margem de segurança e evitar riscos de deficiência (Mazzuco, 2006). Além disso, a utilização de vitaminas como suplemento nas dietas tem evoluído para o atendimento de objetivos específicos, como a melhoria do sistema imunológico e na qualidade dos produtos (Whitehead, 2002). Segundo Leeson & Caston (2003), o enriquecimento de ovos comerciais com vitaminas (até 50% acima dos níveis diários recomendados) atribui uma função nutritiva/funcional à suplementação vitamínica em dietas de poedeiras comerciais.
O aumento continuado na produção de ovos e taxa de crescimento de frangos de corte ditam a necessidade da re-avaliação contínua das especificações de vitaminas dietéticas (Leeson, 2006). De maior significância hoje é a percepção de que níveis dietéticos de vitaminas acima daqueles requeridos para manter objetivos de produção simplistas podem positivamente impactar o valor nutricional de produtos avícolas e também a saúde e bem-estar das aves, como recentemente detalhado em publicações centradas na nutrição vitamínica ótima. As vitaminas atuais podem ser consideradas como nutrientes clássicos, nutracêuticos e moduladores imunes. O futuro provavelmente ditará o conceito de adaptar especificações vitamínicas da dieta para expectativas de produção individual e objetivos que aproveitem as várias funções das vitaminas.
Minerais orgânicos
A avaliação de alimentos e suplementos minerais não depende apenas da concentração ou conteúdo mineral total, mas também de quanto pode ser absorvido no intestino e usado pelas células e tecidos do animal. Isto, em parte, depende da idade e espécie animal; da forma química que o mineral é ingerido; das quantidades e proporções de outros componentes dietéticos com o qual interage metabolicamente (Hazell, 1985; Ammerman et al., 1995, citados por Underwood & Suttler,1999). Assim, Ashmead et al (1985), avaliando as diferentes formas químicas dos minerais no sistema digestório de aves, relataram que os minerais sob a forma de sais sofrem a ação do pH ácido, sendo dissociados e ionizados. Desta forma, esses íons podem ligar-se ou reagir com outras moléculas presentes no trato digestório, como fitatos e oxalatos, tornando-se indisponíveis. No entanto, os pesquisadores verificaram que as moléculas dos minerais na forma orgânica, não sofrem essa dissociação em pH ácido, permanecendo, portanto, com carga elétrica neutra, não podendo assim reagir com outras moléculas presentes na luz intestinal. Logo, os minerais orgânicos são mais bem absorvidos pelas aves e suas concentrações diminuídas nas fezes. A redução da concentração fecal de minerais também foi verificada em suínos (Burkett et al., 2006) suplementados com minerais orgânicos (Bioplex). Os autores relataram a diminuição na concentração fecal de Fe, Cu e Zn sem afetar o desempenho dos animais.
Avaliando os efeitos da suplementação de rações com níveis crescentes de cobre de fonte orgânica, comparado à fonte inorgânica, sobre o desempenho e incidência de diarréia em leitões desmamados, Muniz et al. (2007) não verificaram efeitos da suplementação de cobre nas formas orgânica ou inorgânica sobre a conversão alimentar e a incidência de diarréia nos leitões, bem como diferenças para consumo diário de ração e ganho diário de peso dos leitões para cada nível de cobre na forma orgânica (50, 100 e 150 ppm), em relação ao tratamento com 240 ppm de cobre na forma de sulfato, demonstrando a possibilidade da substituição do cobre inorgânico por níveis bastante inferiores de cobre orgânico nas dietas, sem prejuízos ao desempenho dos leitões em qualquer um dos períodos avaliados. Entretanto, observaram efeito quadrático dos níveis crescentes de cobre sobre o consumo diário de ração, sendo 90 ppm na forma orgânica o melhor nível para estimular consumo.
Selênio
O selênio é um mineral-traço essencial, tanto para humanos quanto para animais. No entanto, o selênio é o mais tóxico de todo os elementos-traço e podem surgir grandes problemas devido à administração inconseqüente de suplementos desse mineral (Underwood & Suttle, 1999). A margem de segurança entre a toxidez e a concentração dietética necessária é bastante estreita e a maioria dos experimentos com animais são conduzidos na forma de dose-resposta, variando entre níveis deficientes, sub-ótimos e ótimos, que variam entre os níveis praticados correntemente num extremo, e a tolerância ponderada, em outro. O selênio é necessário para o crescimento e fertilidade nos animais e para a prevenção de uma série de doenças, podendo apresentar respostas variáveis à vitamina E. Pesquisas com ratos confirmaram que a vitamina E e o selênio atuam sinergicamente (Awad et al., 1994; Levander et al. 1995, citados por Underwood & Suttle, 1999), sem que, no entanto, um nutriente compense completamente a deficiência ao outro.
A maioria das dietas de animais de produção são deficientes em selênio, uma vez que a maioria dos cereais e farelos utilizados na elaboração das rações são pobres nesse mineral. Por muitos anos, tais dietas vêm sendo suplementadas com selênio, principalmente na forma inorgânica, como o selenito de sódio. Esta manobra nutricional tem parcialmente auxiliado na prevenção de síndromes decorrentes da deficiência em selênio, como a diátese exudativa, fibrose pancreática, além de desordens reprodutivas, em aves, e miopatias cardíacas, em suínos (McCartney, 2005). Apesar da importância fisiológica e de seus efeitos na produtividade de aves comerciais, a utilização do selênio inorgânico tem sido relacionada a fatores indesejáveis. As limitações para uso do selênio inorgânico são bem conhecidas: toxidez, interação com outros minerais, baixa eficiência para a produção de leite e carne. Além disso, a baixa retenção corporal do selênio inorgânico leva a deficiência do animal em um espaço de tempo muito curto; por sua vez, o selênio orgânico na forma de selênio-aminoácido forma parte da proteína endógena e como tal não causa risco de deficiência de selênio.
Diante disso, experimentos com aves de postura, mostraram bons resultados na produção de ovos com o uso do Sel-Plex sozinho ou combinado com outros minerais orgânicos (Pan et al., 2004; Xavier et al., 2004). A razão para essa melhora com a suplementação orgânica do selênio pode estar relacionada à selenometionina, que pode ser armazenada no organismo da ave. Cave (1984) e Brake et al. (1985) atribuíram o aumento na produção de ovos em seus experimentos ao aumento na deposição da proteína no organismo. Portanto, estes resultados sugerem que o aumento da produção de ovos esta relacionado ao aumento na quantidade de folículos maduros em resposta ao aumento na síntese de receptores das gonadotrofinas. A proteção antioxidante da enzima GSH-Px e a função do Zn na síntese de proteína podem ter contribuído para a síntese e liberação dos hormônios LH e FSH e síntese dos seus receptores. De acordo com Franco e Sakamoto (2007), a suplementação de selênio nas rações de poedeiras não apenas previne os sintomas de deficiência, mas também permite aumentar a concentração deste mineral nos ovos, o que favorece uma maior ingestão de selênio pelos consumidores deste produto. Além disso, o incremento dos níveis de selênio nos ovos possibilita a manutenção da qualidade interna dos ovos durante os períodos de estocagem.
Nucleotídeos
Os nucleotídeos purina e pirimidina estão envolvidos em praticamente todos os processos celulares e exercem papel fundamental sobre as funções estruturais, metabólicas, energéticas e regulatórias. As fontes de nucleotídeos dos enterócitos são provenientes da suplementação dietética e da síntese de novo. A demanda nutricional para a proliferação e diferenciação de linfócitos na camada mucosa dos intestinos e nos enterócitos é bastante alta. Os nucleotídeos estão naturalmente presentes em todos os alimentos de origem animal ou vegetal na forma de nucleotídeos livres e ácidos nucléicos. As concentrações de nucleotídeos nos alimentos dependem principalmente de sua densidade celular. Geralmente, os grãos possuem um conteúdo de ácidos nucléicos maior, mas o suprimento de nucleotídeos para a utilização pelo animal deve ser hidrolisado no trato digestivo (Lee et al., 2007).
Segundo Rutz (2006), o desenvolvimento de dietas pré-iniciais para aves e suínos e os avanços no conhecimento da fisiologia digestiva e sua relação com a imunidade criaram um novo campo, denominado imuno-nutrição, que busca o fornecimento de bionutrientes ativos que auxiliem no desenvolvimento precoce do trato gastrointestinal e que gerem resultados produtivos melhores. Neste contexto, a suplementação dietética com nucleotídeos é relatada por Córdoba et al. (2008) como uma estratégia para melhorar a saúde e o desempenho de leitões na fase pósdesmame. O desmame geralmente causa um declínio no consumo de ração pelos leitões, que pode acarretar em atrofia das vilosidades intestinais, com conseqüente redução da capacidade absortiva de efeitos na produtividade.
De acordo com Tibbets (2007), os principais benefícios da utilização de nucleotídeos nas dietas para animais estão relacionados à maior eficiência no metabolismo energético e nitrogenado mediante a melhoria da morfologia intestinal e maior absorção, que por sua vez, resulta na melhor reposta imune com a redução de desordens intestinais e em alta taxa de crescimento de tecidos.
Existem diversas fontes de nucleotídeos, entre elas a levedura. O extrato de levedura é rico em nucleotídeos, inositol (importante promotor de crescimento), glutamato (estimulante de palatabilidade), proteínas, vitaminas e minerais (Tibbetts, 2004).
A utilização do extrato de levedura nos estágios iniciais de produção aumenta o crescimento, consumo de ração e eficiência alimentar, além de produzir melhoras na morfologia intestinal e saúde animal, a curto e longo prazo. Segundo EsteveGarcia et al. (2007), durante períodos de intenso desenvolvimento, a disponibilidade de nucleotídeos pode limitar a maturação de tecidos de rápida replicação celular e baixa capacidade biossintética, como o intestino, restringindo, desta forma, a taxa de crescimento, pela menor capacidade de digestão e absorção de nutrientes.
Avaliando o efeito da utilização de um extrato de levedura (NuPro®) sobre o desempenho e características de carcaça de frangos de corte, Rutz et al. (2006) verificaram que o desempenho produtivo foi melhor ao se fornecer extrato de levedura de 1 a 7 e de 38 a 42 dias de idade, embora as carcaças não tenham sido afetadas pelos tratamentos. Os autores atribuíram o melhor desempenho das aves ao aumento da relação vilo:cripta, propiciado por ação de nucleotídeos presentes no extrato de leveduras, favorecendo o aumento da capacidade de digestão e absorção de nutrientes. Citam, ainda, que a presença de inositol pode ter contribuído para o aumento do peso corporal, pois este é um promotor natural do crescimento. Além disso, o inositol está relacionado com fosfolipídios, que são componentes estruturais das membranas que, por sua vez, exercem papel na permeabilidade seletiva e na regulação de receptores hormonais situados na superfície celular (McDowell, 1989).
Probióticos
A utilização de probióticos representa uma alternativa eficiente na substituição dos antibióticos na manutenção de altos índices de produtividade e qualidade dos produtos finais na criação de aves. O uso de probióticos está fundamentado no controle biológico realizado por microorganismos vivos específicos suplementados na dieta com o objetivo de colonizar o trato gastrintestinal com microrganismos benéficos por meio de exclusão competitiva, garantindo melhoria na saúde do animal e maior eficiência no aproveitamento dos nutrientes da ração. Além da exclusão competitiva, também são observados antagonismo direto e melhoria da resposta imune humoral. Além disso, a utilização de probióticos agrega valor ao frango, uma vez que este não deixa resíduos nos produtos de origem animal e não provoca resistência às drogas utilizadas em seres humanos. Estas razões vêm sendo aceitas pelo consumidor, principalmente dos grandes centros urbanos, nos quais se observa uma crescente conscientização das vantagens de consumir carnes com menores riscos da presença de resíduos. Podem-se somar a estas razões os perigos que o uso contínuo dos antimicrobianos nas rações representa no surgimento de resistências múltiplas de bactérias, assim como os impactos negativos que isto pode ter na saúde e no ambiente (Santos et al., 2004).
Flemming & Freitas (2005) conduziram um estudo com o objetivo de comparar o efeito do uso de probiótico (Bacillus licheniformis e Bacillus subtilis), probiótico mais mananoligossacarídeos (MOS), promotor de crescimento (avilamicina) e uma dieta controle sem aditivos sobre o desempenho de frangos de corte. Concluiu-se que na primeira semana as aves alimentadas com rações contendo o probiótico e o MOS apresentaram um ganho de peso melhor que os demais grupos. A conversão alimentar do grupo que recebeu probiótico na primeira semana foi melhor do que a do grupo que recebeu promotor de crescimento, não diferindo da inclusão de MOS ou da dieta controle. No resultado acumulado (início ao abate com 42 dias), o probiótico apresentou os melhores resultados para ganho de peso em relação ao controle, não diferindo, entretanto da associação de probiótico com prebiótico (MOS) e o antibiótico.
Prebióticos
São ingredientes não nutrientes, não digeríveis, que afetam beneficamente o hospedeiro, estimulando seletivamente o crescimento e a atividade de uma ou mais espécies de bactérias benéficas intestinais. Este efeito ocorre principalmente por estimular seletivamente o crescimento e/ou a atividade de número limitado de bactérias nos organismo. Como exemplo, tem-se os mananoligossacarídeos (MOS) – que são extraídos da parede celular de Saccharomyces cerevisiae e contém Dmanose e, os frutoligossacarídeos (FOS) – sintetizados a partir do Aspergillus niger ou Aureobasidium pullulans. Existem ainda os glicoligossacarídeos e os galactoligossacarídeos.
Os prebióticos têm sido apontados como possíveis substitutos aos antibióticos e têm seus efeitos baseados na redução do crescimento de muitas bactérias enteropatogênicas pela redução do pH, resultante do aumento da produção de ácido lático nos cecos. Algumas bactérias podem reconhecer sítios de ligação nas moléculas dos prebióticos, reduzindo a colonização da mucosa intestinal.
De acordo com Furlan et al. (2004), a alteração da microbiota intestinal causada pelo uso de prebióticos pode ocorrer de duas maneiras: através do fornecimento de nutrientes para as bactérias desejáveis ou através do reconhecimento, pelas bactérias patogênicas, de sítios de ligações nos oligossacarídeos como sendo da mucosa intestinal, reduzindo a colonização intestinal indesejável, resultando em menor incidência de infecções e melhor integridade da mucosa intestinal. Os oligossacarídeos no intestino promovem a modificação do ecossistema bacteriano com aumento no número de Bifidobacterium e Lactobacillus, que suprimem a atividade de bactérias putrefativas e reduzem a formação de produtos tóxicos da fermentação, tais como amônia, aminas e nitrosaminas (Flickinger et al., 2003).
Albino et al. (2006) avaliaram o rendimento de cortes e o desempenho de frangos alimentados com dietas contendo mananoligossacarídeos e a viabilidade de substituição dos antibióticos por aditivos alternativos. Foram utilizados três tipos de aditivos: um promotor de crescimento (avilamicina) e dois prebióticos (mananoligossacarídeos) com diferentes concentrações de leveduras, denominados MOS standard e MOS alta concentração, incluídos nas dietas na forma isolada ou associados. Verificou-se que a adição de avilamicina e do mananoligossacarídeo standard, isoladamente ou em combinação, proporcionou efeitos benéficos no ganho de peso das aves no período de 1 a 42 dias de idade. Os autores concluíram que prebióticos à base de mananoligossacarídeo podem substituir o antibiótico avilamicina em rações para frangos de corte sem prejuízos à produção.
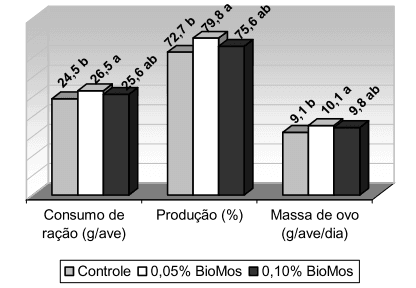
Costa et al. (2008), trabalhando com codornas de postura, verificaram maiores valores de produção de ovo e massa de ovo em aves que receberam dieta contendo 0,05% de Bio-Mos. Porém, quando se aumentou a suplementação de Bio-Mos para 0,10%, o desempenho foi superior ao apresentado pelas aves submetidas à dieta controle, mas inferior ao daquelas que receberam 0,05% de Bio-Mos (Figura 1). Uma provável explicação está no fato das codornas apresentarem taxa de passagem do alimento mais rápida e um maior consumo de ração relativo ao peso corporal, quando comparado às galinhas. A presença da maior quantidade de oligossacarídeos na dieta com 0,10% de suplementação pode ter estimulado ainda mais a velocidade do trânsito intestinal, reduzindo a digestibilidade dos nutrientes em relação à dieta com 0,05% de Bio-Mos.
Figura 1 – Efeito da adição de prebióticos na dieta sobre a produção de ovo e a massa de ovo de codornas japonesas em postura
Uso de adsorventes nas rações para minimizar os efeitos das micotoxinas
Os adsorventes são substâncias capazes de ligar-se a micotoxinas e transportá-las para fora do TGI impedindo a intoxicação por micotoxinas. Entre os adsorventes utilizados na produção animal estão os aluminosilicatos, bentonita e mananoligossacarídeos (Bellaver, 2000).
O efeito da micotoxina do Fusarium e da utilização de adsorvente sobre o desempenho de poedeiras de 0 a 8 semanas de postura revelou que a aflatoxina aumentou o consumo de ração das aves e piorou a conversão alimentar em quilograma de ração por massa de ovo produzida (Smith et al.,2004). No entanto, a adição de adsorvente resultou em conversão alimentar semelhante à das aves do grupo controle (sem aflatoxina). Semelhantemente, Mallmann & Rauber (2006) avaliaram a influência das aflatoxinas adicionadas à dieta e a eficiência de um adsorvente de micotoxinas, sobre o desempenho de perus de corte intoxicados com 500 ppb de aflatoxinas, durante um período de 21 dias. Verificou-se que a aflatoxina apresentou efeito deletério sobre o desenvolvimento de perus de corte durante os primeiros 21 dias de vida. Além disso, pôde-se observar que o adsorvente utilizado apresentou resultados satisfatórios no controle dos efeitos tóxicos destas micotoxinas.
Enzimas
Um grande número de trabalhos realizados no mundo inteiro têm validado a eficiência da suplementação com enzimas exógenas em dietas para aves e suínos. Os efeitos dietéticos das enzimas exógenas estão associados ao aumento da digestão de produtos de baixa qualidade e redução da perda de nutrientes nas fezes, sendo assim possível reduzir os níveis nutricionais da dieta com possíveis vantagens econômicas. Para tal, uma variedade de carboidrases, proteases, fitases e lipases são utilizadas para estes fins (McCleary, 2001). Normalmente, as enzimas comerciais usadas como aditivos são preparados enzimáticos contendo uma variedade de enzimas, uma vez que as rações são compostas por diversos ingredientes (Campestrini et al., 2005). Os polissacarídeos não-amiláceos (PNAs) podem ser potencialmente aproveitados pelo animal mediante a utilização de enzimas exógenas que hidrolisam estes compostos, aumentando o aproveitamento da energia presente nos alimentos. Adicionalmente, a hidrólise dos PNAs promovem a redução da viscosidade da digesta (Slominski et al., 2006; Buchanan et al., 2007).
A utilização da fitase já está bem estabelecida, sendo ampla a literatura a esse respeito. A enzima fitase, hidrolisa o fitato presente em todos os ingredientes de origem vegetal, tornando disponível tanto o fósforo, como outros elementos, indiretamente, tornando-os disponíveis para o metabolismo animal (Roland et al., 2006). O fitato é uma molécula polianiônica com potencial para quelatar nutrientes positivamente carregados (Na+, Mg++, K+, Ca++ e Zn++, entre outros), comprometendo a utilização de proteínas, energia, cálcio e minerais-traço (Selle & Ravindran, 2007).
A disponibilidade e variedade de grãos no Brasil são bastante grandes, o que permite a inclusão e/ou substituição total ou parcial de determinados ingredientes por outros que apresentem preços reduzidos, principalmente em períodos de entressafra. No entanto, tais ingredientes ditos alternativos apresentam restrições quanto ao seu uso na formulação das dietas para aves, por conterem fatores antinutricionais, que prejudicam o desempenho, e conseqüentemente, resultam em baixa uniformidade e lucratividade ao final da produção. O uso de enzimas permite que tais ingredientes sejam utilizados de maneira tão eficiente quanto o são o milho e a soja (Campestrini et al., 2005).
Segundo Wyatt & Bedford (1998), existem diferentes abordagens econômicas ao se considerar a incorporação de enzimas exógenas nas formulações das dietas. Uma aplicação mais simples e provavelmente mais prática, chamada de "over the top" (por cima) com intuito de melhorar o desempenho de forma mais econômica, que consiste em suplementar com as enzimas uma formulação padrão, sem alterar os níveis nutricionais. Uma segunda alternativa seria alterar a formulação da ração reduzindo os nutrientes e adicionando enzimas exógenas para restaurar o valor nutricional da dieta padrão, visando o mesmo desempenho de uma dieta com os níveis nutricionais normais. Se a suplementação for eficiente, os parâmetros produtivos seriam os mesmos (Zanella, 1999). Deve-se avaliar ainda a possibilidade da utilização das enzimas em dietas reformuladas, nas quais a matriz nutricional da enzima deve ser considerada na formulação das dietas, o que possibilitaria uma maior redução nos custos de arraçoamento, haja vista as enzimas contribuírem com cerca de 50 a 75 kcal de EM; 0,1% de Ca e 0,1% de P disponível por quilograma de ração (Wyatt & Bedford, 1998).
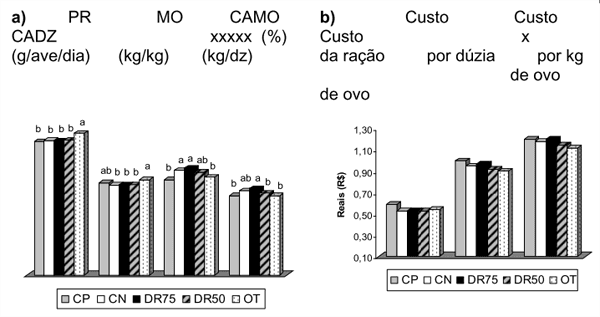
Em experimentos criteriosos, Costa et al. (2008) avaliaram o efeito de enzimas exógenas fibrolíticas sobre o desempenho e qualidade dos ovos de poedeiras em segundo ciclo de produção (Figura 2), onde verificaram que a utilização de enzimas promoveu maior viabilidade econômica, resultando em menor custo por dúzia e por massa de ovo produzido quando comparado aos demais tratamentos (Costa et al., 2008a). Resultados semelhantes confirmaram a eficiência do complexo multienzimático no desempenho de aves em pico de postura (Costa et al., 2008b; Figura 3).
Figura 2 – Efeito da adição das enzimas exógenas em dietas para poedeiras de segundo ciclo de produção sobre o desempenho (a) e custos da ração e do arraçoamento por dúzia e por massa de ovo produzido (b)
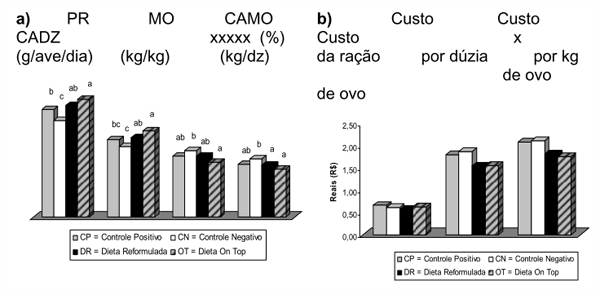
Figura 3 – Efeito da adição das enzimas exógenas em dietas para poedeiras no pico de produção sobre o desempenho (a) e custos da ração e do arraçoamento por dúzia e por massa de ovo produzido (b)
Estudos mostram que a utilização de enzimas xilanolíticas inibe a fermentação no íleo e estimula a fermentação nos cecos (Bedford, 2001; Persia et al., 2002). A redução da fermentação ileal é benéfica para o hospedeiro, pois grande parte do material fermentado nesta região constitui-se de amido e proteína não digeridos, que, assim, ficariam disponíveis para serem hidrolisados e absorvidos pelo hospedeiro (Bedford, 1996). Além disso, os oligossacarídeos resultantes da degradação dos PNAs pelas enzimas exógenas teriam um efeito prebiótico no ceco, atuando como substrato para a proliferação de bactérias benéficas ao hospedeiro em detrimento de bactérias patogênicas, melhorando, desta forma, a saúde da ave (Persia et al., 2002).
A poluição ambiental, pela excreção fecal de nitrogênio e fósforo, pode se dar em maior ou menor grau, dependendo da capacidade de utilização desses nutrientes, pelos animais, que é melhorada com a adição de enzimas exógenas. Suínos e aves excretam mais da metade do fósforo e do nitrogênio que consomem. O uso de enzimas na ração das aves e suínos e outros animais domésticos, melhora a digestibilidade e disponibilidade de certos nutrientes para os animais, principalmente o fósforo, nitrogênio, cálcio, cobre e zinco, diminuindo a sua presença nas fezes e urina, e conseqüentemente, a sua deposição no meio ambiente (Campestrini et al., 2005).
As fitases apresentam aplicações generalizadas, uma vez que seu substrato está presente invariavelmente em dietas para aves e suínos, e sua inclusão resulta em maior biodisponibilidade de fósforo e na redução da excreção deste mineral no ambiente. A proibição do uso de farinhas protéicas de origem animal, que também são fontes de fósforo, tem acelerado a aceitação e utilização de fitase como aditivo em rações para animais em alguns países. A capacidade desta enzima de liberar o fósforo fítico e reduzir a excreção para o meio ambiente está bem documentada; a fitase é uma fonte econômica e alternativa de fósforo e, uma vez que as reservas naturais de fósforo não são renováveis, o seu uso seria benéfico, inclusive, para a preservação de tais contingentes (Selle e Ravindran, 2007).
Estudos realizados por Leytem et al. (2008) e Dozier et al. (2008) demonstram que a redução do fósforo fitato nas dietas pode resultar em menor excreção deste mineral nas excretas das aves. Silva et al. (2008) verificaram que a manipulação dos níveis de proteína e P em rações suplementadas com aminoácidos e fitase pode reduzir, principalmente, a excreção de P, N e Cu, minimizando a poluição ambiental causada por esses elementos.
Portanto, as enzimas exógenas se apresentam como ferramenta na flexibilização das formulações de dietas para aves, possibilitando a utilização de ingredientes não-convencionais sem prejuízo ao desempenho das aves, com conseqüente redução dos custos de produção. Adicionalmente, a suplementação enzimática pode reduzir problemas ambientais, melhorar o ambiente de criação, reduzindo lesões, melhorando, desta forma o bem-estar das aves.
Resíduos secos de destilaria com solúveis (DDGS)
As projeções para os próximos dez anos apontam para a crescente participação do milho no cenário mundial como também já se constatou o aumento exponencial do número de usinas produtoras de etanol norte-americano (USDA, 2007). Contudo, a substituição da fonte energética convencional por fontes renováveis, como o etanol proveniente do milho, elevaram o custo de produção aumentando ainda mais o interesse por ingredientes alternativos. Idéias inovadoras têm surgido em diversas direções entre as quais o uso de resíduos e a utilização de gorduras de baixa digestibilidade, mediante a utilização de fosfolipídios, parecem ser medidas apropriadas em curto prazo.
A maioria dos trabalhos de pesquisa sobre alimentação animal está focalizada no uso de co-produtos e na promoção destes ingredientes a fim de reduzir custos tanto na produção animal quanto na indústria de alimentos e combustíveis (Balakrishnan, 2004). A utilização de resíduos secos de destilaria com solúveis (DDGS) na alimentação de não-ruminantes (Fastinger et al, 2006) é uma proposta relativamente nova, visto o DDGS ser historicamente utilizado na criação de ruminantes devido ao alto conteúdo em fibra (Ham et al., 1994).
O DDGS é um subproduto da fabricação de etanol; são resíduos secos restantes após a fermentação do amido do milho pelas leveduras e enzimas selecionadas para produzir o etanol e o dióxido de carbono. A variabilidade nos níveis nutricionais do DDGS e o baixo valor biológico da proteína são fatores limitantes ao seu uso em dietas para frangos de corte (Fastinger et al, 2006). Além disso, assim como no processamento da soja, o escurecimento do DDGS sugere um aquecimento excessivo e provavelmente a diminuição na digestibilidade de aminoácidos (Parsons et al., 2006). Somado à variação na energia disponível, outro fator considerável é o teor do sódio das amostras de DDGS (0.05 a 0.17%) que é até oito vezes superiores ao do milho (0.02%) (Parsons et al, 2006). No entanto, trabalhos desenvolvidos pelo grupo de pesquisa do Dr. Waldroup da Universidade de Arkansas (EUA) têm encontrado resultados da utilização em dietas para frangos de corte em ate 15 a 20% de DDGS sem afetar o desempenho das aves (Wang et al, 2007).
Para suínos, a inclusão recomendada é de até 30% da dieta, porém, para porcas em gestação, existem estudos que sugerem inclusão de 50%. Foi demonstrado que para porcas, 15 a 30% de DDGS na dieta não afetaram o desempenho nem das porcas nem da leitegada (Leonel & Sousa, 2007). Segundo Brito (2008), a inclusão de 10% de DDGS tem sido a mais recomendada, pois, não se observa diferença no desempenho de suínos alimentados com dieta à base de milho e farelo de soja. O autor ressalva que inclusões de 20 e 30% podem reduzir alguns parâmetros de desempenho o qual pode ser devido à impossibilidade de encontrar a exigência dos aminoácidos digestíveis (Brito, 2008). Para frangos de corte os valores de inclusão de DDGS são inferiores aos utilizados para suínos, segundo Scheideler (2005) a maioria dos nutricionistas utiliza de 8 a 10%, aparentemente pela maior capacidade digestiva de suínos em utilizar ingredientes moderadamente fibrosos.
Para aves poedeiras, a inclusão de 30% de DDGS na dieta não afetou o número de ovos/ave/ano, sendo que, existem trabalhos onde 10% de DDGS na dieta não interferem na produção, no peso dos ovos e nem na qualidade da casca. Alguns autores sugerem que o DDGS tem efeito positivo sobre a qualidade interna do ovo (aumento na unidade Haugh) quando comparado com outras fontes protéicas como farelo de soja e trigo (Leonel & Sousa, 2007).
Recomendações de inclusão de DDGS acima desse valor estão freqüentemente associadas à adição de enzimas fibrolíticas (Gaines et al., 2007 e Spencer et al., 2007 citados por Holtkamp et al, 2007). A adição de fitase juntamente com xilanase em uma dieta para suínos à base de milho e farelo de soja com inclusão de 20% de DDGS resultou em aumento de 5,6% na digestibilidade da energia (175 kcal/kg) e aproximadamente 4,5% na digestibilidade ileal dos aminoácidos. A digestibilidade do fósforo aumentou de 22 para 51% (Barletta & Cooksley, 2008).
Glicerina
Outro subproduto resultante de produção de biocombustíveis, tanto a partir do milho quanto da gordura de frangos, a glicerina, é um potencial substituto parcial do milho para alimentação animal (Cerrate et al., 2006). A glicerina foi eficientemente utilizada por frangos de corte cujo valor médio assemelhou-se ao do milho (3.434 kcal/kg) (Dozier et al., 2008). A adição de glicerina até o nível de 5% não mostrou nenhum efeito adverso no crescimento ou no rendimento da carcaça. O uso de 10% de glicerina resultou na redução do desempenho dos frangos, que foi grandemente relacionado a problemas na taxa de passagem (Cerrate et al., 2006).
Fosfolipídeos
Óleos e gorduras são as mais importantes e caras fontes de energia (Blanch et al., 1996). Além disso, após a elevação substancial no preço do grão devido ao deslocamento da produção na geração de energia para outros setores tecnológicos, a busca da melhoria pela utilização da gordura animal dietética transformou-se em interesse mundial. De acordo com o entendimento corrente, a suplementação dietética com fosfolipídios aumenta substancialmente a digestibilidade de ácidos graxos saturados, diminuindo efetivamente o custo da ração. A recomendação do uso de gorduras em dietas típicas para aves e suínos em países asiáticos tem sugerido que a suplementação de 1 a 4 kg/MT de fosfolipídios promove a melhoria na utilização de nutrientes e desempenho animal (Mu, 2007). Dessa maneira, a inclusão de fosfolipídios dietéticos bem como seus efeitos tem sido investigada em diversas direções (Huang et al, 2008).
As mudanças na disponibilidade e no preço dos principais ingredientes que fornecem energia às dietas resultarão em mudanças nos padrões de formulação (Penz Júnior & Gianfellici, 2007). De uma maneira geral, de acordo com essas recomendações, os nutricionistas podem ajustar a especificação das formulações para reduzir os custos de alimentação, mantendo ou melhorando o desempenho animal.
Pode-se discutir qual seria a alternativa mais interessante na formulação de dietas comerciais, porém não restam dúvidas que o uso do DDGS, glicerina e fosfolipídios em dietas animais são propostas comerciais de um mundo globalizado e sem fronteiras.
Considerações finais
A biotecnologia torna-se ainda mais importante para um país como o Brasil, que ocupa uma posição privilegiada na produção e exportação de carnes de aves e suínos no competitivo agronegócio mundial, em que as margens são estreitas e os mercados distorcidos por subsídios dos países ricos. Ciências como a Glicômica, com o estudo das estruturas dos açucares para desenvolvimento de prebióticos e adsorventes, e a Nutrigenômica, com o estudo dos efeitos da nutrição na expressão gênica estão sendo as aliadas da nutrição animal, a exemplo do que ocorreu e ocorre com a nutrição humana. A ciência da Nutrigenômica utiliza uma base molecular para explicar como a nutrição pode impactar a saúde, pela alteração da estrutura ou expressão de genes individuais. Conforme esta área avança, métodos que detectam fatores dietéticos que podem impactar a produção animal em nível celular serão caracterizados, os quais, por sua vez, melhorarão o desempenho animal.
Literatura citada
AFTAB, U.; ASHRAF, M.; JIANG, Z. Low protein diets for broilers. World´s Poultry Science Journal, v.62, p.688-701, 2006.
AJINOMOTO [2007] Impacto da disponibilidade de aminoácidos industriais sobre as formulações de rações na américa latina. Disponível em : Acesso em 08/04/08.
ALBINO, L.F.T.; FERES, F.A; DIONIZIO, M.A. et al. Uso de prebióticos à base de mananoligossacarídeo em rações para frangos de corte. Revista Brasileira de Zootecnia, v.35, n.3, p.742-749, 2006.
AMMERMAN, C.B., HENRY, P.R.; MILES, D. Supplemental organically-bound mineral compounds in livestock nutrition. In: GARNSWORTHY, P.C.; WISEMAN, J. (Eds), Recent Advances in Animal Nutrition, Nottingham University. 1998
ARAÚJO, L.F.; JUNQUEIRA, O.M.; ARAÚJO, C.S.S. Redução do nível protéico da dieta, através da formulação baseada em aminoácidos digestíveis. Ciência Rural, Santa Maria, v.34, n 4, p.1197-1201, 2004.
ASHMEAD, H.D.; GRAFF, D.J.; ASHMEAD, H. H. Intestinal absorption of metal ions and chelates. Springfield: Charles C Thomas, 1985. p.13-26.
AVICULTURA INDUSTRIAL. [2008] Abate de frangos chega a 1,1 BI de cabeças no 4o trimestre de 2007. Disponível em: Acesso em: 08/04/08.
BALAKRISHNAN, V. Developments in the Indian feed and poultry industry and formulation of rations based on local resources. In: PROTEIN SOURCES FOR THE ANIMAL FEED INDUSTRY EXPERT CONSULTATION AND WORKSHOP. 2004, Bangkok. Proceedings…Bangkok:FAO Animal Production and Health Proceedings, 2004. 390 p.
BARLETTA, A.; COOKSLEY, J. [2008] Enzyme technology significantly improves digestibility of DDGS. Disponível em : Acesso em: 08/04/2008.
BEDFORD, M. R. Enzymes, antibiotics and intestinal microflora. Feed Mix, v.9, n.2, 2001.
BELLAVER, C. O uso de microingredientes (aditivos) na formulação de dietas para suínos e suas implicações na produção e na segurança alimentar. In: CONGRESSO MERCOSUR DE PRODUCCIÓN PORCINA, 2000, Buenos Aires. Anais… Buenos Aires: Embrapa CNPSA, 2000.
BLANCH, A.; BARROETA, A.; BAUCELLS, M. Utilization of different fats and oils by adult chickens as a source of energy, lipid and fatty acids. Animal Feed Science and Technology, v.61, p. 335–342, 1996.
BRAKE, J.; GARLICH, J.D.; PEEBLES, E.D. Effect of protein and energy intake by broiler breeders during the prebreeder transition period on subsequent reproductive performance. Poultry Science, v. 64, p.2335-2340, 1985.
BRITO, C. [2008] Uso do DDGS, um subproduto na produção do etanol, na alimentação de monogástricos. Disponível em: Acesso em 08/04/2008. BUCHANAN, N.P.; KIMBLER, L.B.; PARSONS, A.S. et al. The effects of nonstarch polysaccharide enzyme addition and dietary energy restriction on performance and carcass quality of organic broiler chickens. Journal of Applied Poultry Research, v.16, p.1–12, 2007.
CAMPESTRINI, E.; SILVA, V. T. M da; APPELT, M. D. Utilização de enzimas na alimentação animal. Revista Eletrônica Nutritime, v.2, n.6, p.254-267, 2005.
CAVE, N.A.G. Effect of a high-protein diet fed prior to the onset of lay on performance of broiler breeder pullets. Poultry Science. v.63, p.1823-1827, 1984.
CERRATE S, Y.A.N.; WANG, F.; COTO, Z. et al. Evaluation of glycerine from biodiesel production as a feed ingredient for broilers. International Journal of Poultry Science v.5, p.1001-1007, 2006.
CÓRDOBA, X.; BORDA, E.; MARTÍNEZ-PUIG, D. et al. The role of nucleotides in piglet nutrition. Pig Progress. v.24, n.1, 2008.
COSTA, F.G.P.; NOBRE, I.S., GOULART, C.C. et al. Utilization of prebiotic in laying quails (Coturnix coturnix japonica) diet. In: ALLTECH INTERNATIONAL ANIMAL HEALTH AND NUTRITION SYMPOSIUM, 24. 2008. Lexington. Proceedings… Lexington: Alltech, 2008. Cd-rom.
COSTA, F.G.P.; ROSTAGNO, H.S.; ALBINO, L.F.T. et al. Níveis dietéticos de proteína bruta para frangos de corte de 1 a 21 e 22 a 42 dias de idade. Revista Brasileira de Zootecnia, v.30, n.5, p.1498-1515, 2001.
DEAN, D.W.; BIDNER, T.D.; SOUTHERN, L.L. Glycine supplementation to low protein, amino acid-supplemented diets supports optimal performance of broiler chicks. Poultry Science, v.85, p.288–296, 2006.
HOLTKAMP, D.; ROTTO, H.; GARCIA, R. [2007] The economic cost of major health challenges in large US swine production systems. Disponível em:http://mark.asci.ncsu.edu/Swine_News/2007/sn_v 3004%20 (May).htm> Acesso em: 28/03/2008.
DOZIER III, W.A.; KIDD, M.T.; CORZO, A. Live performance and environmental impact of broiler chickens fed diets varying in amino acids and phytase. Animal Feed Science and Technology, v.141, p.92–103, 2008.
DOZIER, W. A., III, KERR, B. J., CORZO, A., KIDD, M. T., WEBER, T. E., BREGENDAHL,K. Apparent Metabolizable Energy of Glycerin for Broiler Chickens. Poultry Science, v.87, p.317-322, 2008.
EDENS, F.W. An alternative for antibiotics use in poultry: probiotics. Brazilian Journal of Poultry Science, v.5, n.2, p.75-97, 2003.
ESTEVE-GARCIA, E.; MARTÍNEZ-PUIG, D.; BORDA et al. Efficacy of a nucleotide preparation in broiler chickens In: EUROPEAN SYMPOSIUM ON POULTRY NUTRITION, 2007, Strasbourg. Proceedings… Strasbourg:World´s Poultry Science Association, 2007.
ETHERTON, T.D. Melhorias na agricultura animal por meio da biotecnologia. Revista Eletrônica do Departamento de Estado dos EUA, v.. 8, n.3, 2003.
FASTINGER, N.D.; LATSHAW, J.D.; MAHAN, D.C. Amino acid availability and true metabolizable energy content of corn distillers dried grains with solubles in adult cecectomized roosters. Poultry Science, v.85, p.1212-1216, 2006.
FLICKINGER, E. A.; van LOO, J.; FAHEY Jr, G. C. Nutritional responses to the presence of inulin and oligofructose in the diets of domesticated animals: a review. Critical Reviews in Food Science and Nutrition, v.43, n. 1, p.19-60, 2003.
FRANCO , J.R.G.; SAKAMOTO, M.I. [2007] Qualidade dos ovos: uma visão geral dos fatores que a influenciam. Disponível em: Acesso em: 22/03/2008.
FURLAN, R.L.; MACARI, M.; LUQUETTI, B.C. Como avaliar os efeitos do uso de prebióticos, probióticos e flora de exclusão competitiva. In: SIMPÓSIO TÉCNICO DE INCUBAÇÃO, MATRIZES DE CORTE E NUTRIÇÃO, 5. 2004. Balneário Camboriú. Anais... Balneário Camboriú: Fundação Apinco de Ciência e Tecnologias Avícolas, 2004.
GROLLI, D. Agronegócio brasileiro. In: SEMINÁRIO DE AVES E SUÍNOS – AVESUI REGIÕES, 7. 2007, Belo Horizonte. Anais...Belo Horizonte: AveSui, 2007. p.4973.
HAESE, D.; SILVA, B.A.N. Antibióticos como promotores de crescimento em monogástricos. Revista Eletrônica Nutritime, v.1, n.1, p.7-19, 2004.
HAM, G.A.; STOCK, R.A.; KLOPFENSTEIN, T.J. et al. Wet corn distillers byproducts compared with dried corn distillers grains with solubles as a source of protein and energy for ruminants. Journal of Animal Science, v. 77, p.3246– 3257, 1994.
HARVEY,L. Mineral bioavailability. Nutrition and Food Science. v.31, n.4, p.179182, 2001.
HUANG, J., YANG, D., GAO, S. et al. Effects of soy-lecithin on lipid metabolism and hepatic expression of lipogenic genes in broiler chickens. Livestock Science, In Press, Corrected Proof, Available, 2008.
BURKETT, J.L.; STALDER, K.J.; POWERS, W.J. et al. Effect of inorganic, organic, and no trace mineral supplementation on growth performance, fecal excretion, and apparent digestibility of grow-finish pigs In: ALLTECH´S ANNUAL SYMPOSIUM, 22., 2006, Nottingham. Proceedings… Nottingham: Nutritional Biotechnology In The Feed And Food Industries, 2006.
JONDREVILLE, C.; RÉVY, P.S. An update on use of organic minerals in swine nutrition. In: EASTERN NUTRITION CONFERENCE, 2003, Québec. Proceedings… Québec:Animal Nutrition Association of Canada , 2003. p.143158.
KERR, B.J., EASTER, R.A. Effect of feeding reduced protein, amino acidsupplemented diets on nitrogen and energy balance in grower pigs. Journal of Animal Science, v. 73, p. 3000-3008, 1995.
KIDD, M.I.; KERR, B.J.; ALLARD, J.F. et al. Limiting amino acid responses in commercial broilers. Journal Applied Poultry Research, v.9, p.223-233, 2000.
KIDD, M.T.; BURNHAM, D.J.; KERR, B.J. Dietary isoleucine responses in male broiler chickens. British Poultry Science, v.45, n.1, p.67–75, 2004.
KLASING, K. C. Comparative avian nutrition. UK: Cab International, 1998. 350 p.
KOPINSKI, J. [2005] Vitamins for pigs. Disponível em: Acesso em: 28/03/2008
LEE, D. N.; LIU, S. R.; CHEN, Y. T.; WANG, R. C.; LIN, S. Y.; WENG, C. F. Effects of diets supplemented with organic acids and nucleotides on growth, immune responses and digestive tract development in weaned pigs. Journal of Animal Physiology and Animal Nutrition, v. 91, p. 508–518, 2007.
LEESON, S. Vitamin requirements: is there basis for re-evaluating dietary specifications? World´s Poultry Science Journal, v. 63, p. 255-266, 2007.
LEESON, S.; CASTON, L.J. Vitamin Enrichment of Eggs. Journal of Applied Poultry Research, v.12, p.24-26, 2003.
LEONEL, F.P.; D.P. SOUSA. Co-produtos do etanol na alimentação animal. Boletim Técnico Serrana Nutrição Animal, v.79, 2007.
LEYTEM, A.B.; PLUMSTEAD, P.W.; MAGUIRE, R.O. et al. Interaction of calcium and phytate in broiler diets. 2. Effects on total and soluble phosphorus excretion. Poultry Science, v.87, p.459-467, 2008.
MALLMANN, C.A.; RAUBER, R.H.; DILKIN, P. et al. Desempenho produtivo de perus de corte intoxicados com diferentes concentrações de aflatoxinas durante 21 DIAS. Revista Brasileira de Ciência Avícola, v. 8, p. 187, 2006.
MAZZUCO, H. [2006] Vitaminas: funções metabólicas e exigências nutricionais para poedeiras comerciais. Disponível em: Acesso 28/03/2008.
McCLEARY, B.V. Analysis of Feed Enzymes. In: Enzymes in Farm Animal Nutrition, eds M.R. Bedford and G.G. Partridge. CAB International, 2001. 406 p.
McCARTNEY, E. Living longer, happier, healthier? Selenium supplementation of huma foods - the needs to move forward. In: Nutritional biotechnology n the feed and food industries. Proceedings of Alltech´s Twenty First Annual Symposium. Eds. Lyons, T.P. and Jacques, K.A. Nottingam Univesity Press, 2005. 462 p.
McDOWELL, L.R. Vitamins in animal nutrition. San Diego, USA:. Academic Press Inc., 1989. p.486.
MILES, R.D.; HENRY, P.R. Relative trace mineral bioavailability. Ciência Animal Brasileira, Goiânia, v.1, n.2, p.73-93, 2000.
MU, Y. Phospholipids: pratical approach to improving feed value. Magazine Feed International. v.28, n.10, p.18-21, 2007.
MUNIZ, M. H. B. ; BERTO, D. A. ; HAUPTLI, Lucélia ; SILVA, A. M. R. ; TRINDADE NETO, M. A. Cobre de fontes orgânicas e inorgânicas como promotor de crescimento para leitões desmamados. In: CONGRESSO DA ABRAVES, 13., 2007, Florianópolis. Anais… Florianópolis: Abraves, 2007.
NATIONAL RESEARCH COUNCIL - NRC. Nutrient requirement of poultry. 9. ed. Washington, D.C.: National Academic Press,1994. 155p.
NETO, J. P. et. al. Antimicrobianos como aditivos em animais de produção. Farmacologia veterinária. Rio de Janeiro:Editora Guanabara, p. 558- 575, 2002.
PAN, E.A., F. RUTZ, N.J.L. DIONELLO, et al.Performance of brown egg layers fed diets containing organic selenium (Sel-Plex®). In: Annual Symposium Nutritional Biotechnology in the Feed and Food Industries, 20., 2004, Lexington. Proceedings … Lexington, p. 19, 2004.
PARSONS, C. M.; MARTINEZ, C.; SINGH, V.; RADHAKRISHMAN,S.; NOLL, S. Nutritional value of conventional and modified DDGS for poultry. In: MULTISTATE POULTRY NUTRITION AND FEEDING CONFERENCE, 2006, Indianapolis. Proceedings… Indianápolis, 2006.
PEGANOVA, S.; EDER, K. Studies on requirement and excess of isoleucine in laying hens. Poultry Science, v.81, p.1714-1721, 2002.
PELICANO, E.R.L.; SOUZA, P.A.; SOUZA, H.B.A. et al. Desempenho zootécnico de frangosa de corte alimentados com dietas contendo diferentes probióticos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 40., 2003, Santa Maria. Anais ... Santa Maria: Sociedade Brasileira de Zootecnia. Cd-rom.
PENZ JUNIOR, A.M., GIANFELLICI,M. Desafíos de la Nutricion de Pollos de engorda frente a la Competencia de Fuentes Convencionales de energia para la Produccion de Etanol y Biodieses. In: CONGRESO LATINOAMERICANO DE AVICULTURA, 20., 2007, Porto Alegre. Anais… Porto Alegre, 2007, p159-166.
PERSIA, M.E.; DEHORITY, B.A.; LILBURN, M.S. The effects of enzyme supplementation of corn- and wheat-based diets on nutrient digestion and cecal microbial populations in turkeys. Journal of Applied Poultry Research, v.11, p.134–145, 2002.
PLUMSTEAD, P.W.; LEYTEM, A.B.; MAGUIRE, R.O. et al. Interaction of calcium and phytate in broiler diets. 1. Effects on apparent prececal digestibility and retention of phosphorus. Poultry Science, v.87, p.449-458, 2008.
ROLAND, D. A. et al. Comparison of Nathuphos and Phyzyme as phytase sources for commercial layers fed corn-soy diet. In: Poultry Science Association, Inc. 2006.
RUTZ, F.; ANCIUTI, M.A.; RECH, J.L. Desempenho e caracteristicas de carcaças de frangos de corte recebendo extrato de leveduras na dieta. Ciência Animal Brasileira, v. 7, n. 4, p. 349-355, 2006.
SABINO, E.F.N.; SAKOMURA, N.K.; NEME, R. et al. Níveis protéicos na ração de frangos de corte na fase de crescimento. Pesquisa Agropecuária Brasileira, v..39, n.5, 2004.
SANTOS, I.I.; POLI, A.; PADILHA, M.T.S. Desempenho zootécnico e rendimento de carcaça de frangos de corte suplementados com diferentes probióticos e antimicrobianos. Acta Scientiarum. Animal Sciences, v. 26, n. 1, p. 29-33, 2004.
SELLE, P. H; RAVIDRAN, V. Microbial phytase in poultry nutrition: Review. Animal Feed Science and Technology, In. 2007.
SILVA, Y.L.; RODRIGUES, P.B.; FREITAS, R.T.F. et al. Níveis de proteína e fósforo em rações com fitase para frangos de corte, na fase de 14 a 21 dias de idade. 2. Valores energéticos e digestibilidade de nutrientes. Revista Brasileira de Zootecnia, v.37, n.3, p.469-477, 2008.
SLOMINSKI, B. A.; MENG, X.; CAMPBELL, L. D.; GUENTER, W. and JONEST, O. The use of enzyme technology for improved energy utilization from full-fat oilseeds. Part II: Flaxseed, Poultry Science, v. 85, p.1031-1037, 2006.
SMITH T. K.,et al. (2004) Comparative aspects of Fusarium Micotoxicoses in broiler chickens, laying hens and turkeys and the efficacy of a polymeric glucamannan mycotoxin adsobent: Mycosorb. In: ALLTECH´S ANNUAL SYMPOSIUM, 20., 2004. Proceedings... Nutritional Biotechnology in the Feed and Food Industries.p. 103-109.
SUIDA, D. Formulação por proteína ideal e conseqüências técnicas, econômicas e ambientais. In: SIMPÓSIO INTERNACIONAL DE NUTRIÇÃO ANIMAL: PROTEÍNA IDEAL, ENERGIA LIQUIDA E MODELAGEM, 2001, Santa Maria. Anais... Santa Maria: EMBRAPA, 2001. p.27-43.
SUINOCULTURA INDUSTRIAL. [2008] Produção de suínos cresce 6,5% em 2007. Disponível em: Acesso em: 28/03/08
SURAI, P.F. Selenium in nutrition and health. 1st ed. United Kingdom: Nottingham University Press, 2006. 974p.
TALAMINI, D.J.D. Evolução recente e perspectivas da suinocultura brasileira para 2005.
THORNTON, S.A.; CORZO, A.; PHARR, G.T. et al. Valine requirements for immune and growth responses in broilers from 3 to 6 weeks of age. British Poultry Science, v.47, n.2, p.190-199, 2006.
TIBBETTS, G.W. Nucleotídeos presentes no extrato de levedura de cepa específica: alternativa para substituição de fontes protéicas de origem animal. Pork World, p.36-39, 2004.
TOLEDO, J.S.; KLOECKNER, P.; LOPES, J. et al. Níveis das vitaminas A e E em dietas de frangos de corte de 1 a 42 dias de idade. Ciência Rural, v.36, n.2, p.624-629, 2006.
UNDERWOOD, E.J., SUTTLE, N.F. Selenium. In: The mineral nutrition of livestock. 3rd ed. CAB Intrenational. 1999.
WANG, Z., CERRATE, S., COTO, C. et al. Utilization of distillers dried grains with solubles (DDGS) in broiler diets using a standardized nutrient matrix1. International Journal Poultry Science, v.6, n.7, p.470-477, 2007.
WHITEHEAD, C. C. Vitamins in Feedstuffs In: Poultry Feedstuffs: supply, composition, and nutritive value, ed. J. McNab and N. Boorman. Poultry Science Symposium Series. v. 26, 2002. pp.181-190.
WYATT, C.L.; BEDFORD, M. O uso de enzimas nutricionais para maximizar a utilização de nutrientes pelo frango de corte em dietas à base de milho: Recentes processos no desenvolvimento e aplicação prática. In: SEMINÁRIO TÉCNICO FINNFEEDS, 1998, Curitiba. Anais... Curitiba: FINNFEEDS, 1998, p.2-12.
XAVIER, G.B., F. RUTZ, N.J.L. DIONELLO, et al. Performance of layers fed diets containing organic selenium, zinc and manganese, during a second cycle of production. In: ANNUAL SYMPOSIUM NUTRITIONAL BIOTECHNOLOGY IN THE FEED AND FOOD INDUSTRIES, 20., 2004, Lexington. Proceedings … Lexington, p. 19, 2004.
ZANELLA, I.; SAKOMURA, N.K.; SILVERSIDES, F.G. et al. Effect of supplementation of broiler diets based on corn and soybeans. Poultry Science, v.78, p.561-568, 1999.
Tópicos relacionados
Autores:

Junte-se para comentar.
Uma vez que se junte ao Engormix, você poderá participar de todos os conteúdos e fóruns.
* Dados obrigatórios
Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.
Criar uma publicação
26 de abril de 2021
Oferecer uma Alimentação diferenciada para Galinhas Poedeiras, traz resultados para a produçao? (qualidade raçãoXcusto benefício).





Você pode estar interessado em
.jpg&w=3840&q=75)
Evonik Animal Nutrition