INTRODUÇÃO
O melhoramento genético, a competitividade globalizada e o custo dos ingredientes são os pontos básicos que estimulam os pesquisadores a desenvolver modelos e aplicações para reduzir o custo de produção dos frangos de corte. A ração é sem dúvida o ponto crucial a ser investigado, pois representa o maior custo da produção das aves, sendo a energia, a proteína e o fósforo, os elementos que mais influenciam o custo final da dieta, respectivamente.
A redução da proteína bruta da dieta com o uso dos aminoácidos industriais é uma estratégia nutricional que reduz o custo das rações sem comprometer o desempenho final dos frangos de corte. Dentro deste mesmo entendimento, a enzima fitase atuando sobre o fitato reduz o uso do fosfato bicálcico nas rações. Por fim, a valorização da digestibilidade do amido e dos polissacarídeos não amiláceos solúveis dada pelas enzimas carboidrases é outra ferramenta nutricional utilizada para aprimorar a eficiência energética das rações de frangos de corte.
As enzimas carboidrases têm sido amplamente difundidas na alimentação de frangos de corte com o objetivo de valorizar a energia da dieta, aumentando a disponibilidade de glicose ou reduzindo a viscosidade intestinal (Dourado, 2008 e Grecco, 2016). Contudo, a eficiência energética de uma dieta parece ter sua maior dependência do quanto o corpo é capaz de sintetizar o ATP a partir de carboidratos, lipídeos e proteínas (Fernandes e Toro-Velasquez, 2014).
É sabido que os carboidratos e os ácidos graxos, quando absorvidos no intestino, são metabolizados por uma séria de reações de oxidação, produzindo CO2 e H2O. Os intermediários destas reações doam elétrons para as coenzimas nicotinamida adenina dinucleotídeos (NAD+) e flavina adenina dinucleotídeos (FAD) para formar coenzimas reduzidas ricas em energia, NADH e FADH2. Estas coenzimas reduzidas doam um par de elétrons a um conjunto especializado de transportadores de elétrons que perde energia, porém, parte desta energia é capturada e armazenada na forma de ATP a partir do ADP + fosfato inorgânico (Pi). Esse processo é denominado de fosforilação oxidativa. O restante da energia livre não capturada como ATP é liberado na forma de calor (Champe e Harvey, 1997 e Bertechini, 2012).
Como pode ser observado, o ciclo do ácido cítrico atua como principal rota da oxidação dos carboidratos e lipídeos, produzindo ATP e GTP, ou seja, nucleotídeos ligados por três moléculas de fósforo. Segundo Fernandes e Toro-Velasquez (2014), o conteúdo de energia do GTP é 0,75 do ATP. Os dois nucleotídeos são interconversíveis pela reação da enzima nucleosídeo difosfato quinase.
A quantidade de ATP no interior da célula é estritamente regulada e está disponível por um curto período de tempo, caracterizado pela alta demanda de energia pelo corpo, sendo a creatina, um importante elemento que mantém o equilíbrio energético dentro das células musculares (Champe e Harvey, 1997). Porém, esta reserva energética é considerada efêmera, sendo utilizada quando os músculos precisam do máximo esforço. A maior regeneração do ATP é originado pela via anaeróbia, ou seja, a partir da utilização do glicogênio e da glicose até ácido lático, e, também da via aeróbica, do uso da glicose, ácidos graxos e proteínas, produzindo CO2 e H2O.
Conforme supracitado, o metabolismo energético é complexo. O conceito de que o ATP, um nucleotídeo adenina ligado a três moléculas de fósforo, é a fonte universal de energia para as células, determina a hipótese de que qualquer elemento que facilite a sua síntese ou que favoreça a sua disponibilização dentro da célula poderá afetar diretamente o balanço energético da dieta.
Pesquisas com nucleotídeos geralmente foram direcionadas para conhecer o seu efeito no status de saúde animal. Carver e Walker (1995), Koeppel (2000), Mateo (2005) e Hoffmann (2009) mostraram resultados positivos dos nucleotídeos sobre o sistema imune dos animais.
Os nucleotídeos purinas; inosina monofosfato (IMP), guanina monofosfato (GMP) e adenina monofostado (AMP) são considerados intermediários energéticos no organismo dos animais. Dentre os nucleotídeos purinas observados para o processo de fosforilação oxidativa, o IMP parece ser o mais indicado, pois é o precursor do AMP e GMP que são aceptores de grupos fosfatos para formar ATP e GTP os quais ocupam posições intermediárias na escala bioenergética dos compostos fosfatados (Champe e Harvey, 1997).
Atualmente o IMP é utilizado como realçador de sabor, apresentando importante papel no paladar umami (Santos et al, 2015; Mateo, 2005; Loliger, 2000). Não é, portanto, considerado um precursor bioenergético que regenera a energia molecular na forma de ATP (Monteiro, 2015).
As informações referentes ao metabolismo energético deixam evidências de que a suplementação de IMP nas rações, além de seu papel como realçador de sabor, pode ser um elemento precursor de ATP e GTP no organismo animal, criando a expectativa de uma possível fonte de energia para as aves.
O objetivo deste artigo foi investigar o IMP como uma possível fonte complementar de energia na alimentação de frangos de corte.
Inosina Monofosfato como Fonte de Energia para Frangos de Corte
A energia não é quimicamente identificada como um nutriente, mas uma propriedade obtida quando os carboidratos, lipídeos e proteínas são oxidados durante o metabolismo (Klasing, 1998). Quando consumidos pelos animais podem ser usados em três diferentes caminhos: (1) energia suprida para o trabalho; (2) energia convertida em calor e (3) energia estocada no corpo como tecido. Por fim, o consumo excessivo de nutrientes é convertido em gordura que não pode ser excretado do corpo. A utilização ótima dos nutrientes pelas aves é atingida quando a dieta contém uma proporção de energia necessária para os nutrientes atuarem no crescimento, produção de ovos e deposição de proteína corporal. A maior porção de todo o alimento consumido por um animal é usado como energia para atender as reações anabólicas e catabólicas (Scott et al. 1982 e Lesson & Summers, 2001).
A energia metabolizável (EM) é a recomendação comumente usada para as aves. No metabolismo são previstas perdas de energia baseadas na ineficiência do metabolismo intermediário devido à digestão, absorção, transporte, excreção ou transformação de nutrientes em metabólitos ineficientes. Esta perda é referenciada como incremento calórico (Klasing, 1998) que servirá para a manutenção da homeotermia corporal (Bertechini, 2012). O incremento calórico varia de acordo com a composição da dieta e, nas aves, corresponde aproximadamente em 30% da EM para a proteína, 15% de EM para o amido e 10% da EM para os lipídeos. O incremento calórico é menor nos lipídeos por exigirem uma pequena atividade enzimática para a sua digestão, pouca energia requerida para a sua oxidação e por não haver síntese de metabólitos ineficientes. Contrariamente, o incremento calórico para a proteína é alto, pois há intensa atividade enzimática para a sua digestão, alto custo para a sua síntese e necessidade de energia para a excreção de seu metabólito quando os aminoácidos são oxidados para fornecer energia (Klasing, 1998).
Independente disso, o carbono e o hidrogênio presentes nos alimentos podem ser oxidados à dióxido de carbono e água gerando adenosina trifosfato (ATP) e guanosina trifosfato (GTP), que são formados pela união de uma molécula de ácido fosfórico com uma molécula de adenosina difosfato (ADP) e guanosina difosfato (GDP), respectivamente. O aprisionamento da energia via metabolismo da glicose e gordura não somente reduz a perda de calor, mas fornece um constante suprimento de energia para as células do corpo (Scott et al., 1982 e Leeson & Summers, 2001). Em aves, a reação metabólica que oxida os nutrientes captura ao redor de 40% da energia como ATP (Klasing, 1998) e quando o ATP perde um grupo fosfato formando o ADP, há liberação de aproximadamente 8 kcal/mol (Leeson & Summers, 2001).
Para Bertechini (2012) o organismo animal possui uma eficiência energética de aproximadamente 40%. Nas aves e suínos que são animais de sangue quente, a perda de energia do metabolismo na forma de calor (58,8%) é importante na homeotermia.
Os nutricionistas têm os alimentos como única fonte de energia para os animais devido ao conhecimento da digestibilidade e do valor energético dos carboidratos, gorduras e proteínas. Por outro lado, estabelecer a presença dos nucleotídeos purinas como uma possível fonte de energia na nutrição animal não têm sido reconhecido no meio científico e nas indústrias de rações e de aves.
Por mais de duas décadas, os nucleotídeos têm sido estudados mediante ao fato de participarem de várias reações fisiológicas que são essenciais para a manutenção e propagação da vida (Mateo, 2005). Os nucleotídeos estão envolvidos em quase todos os processos que determinam o papel estrutural, metabólico, energético e funções regulatórias. Eles compõem as unidades monoméricas do RNA e DNA; o RNA é requerido para a síntese de proteína e o DNA é necessário para o crescimento e divisão celular. O ATP, um nucleotídeo adenina, é a maior fonte de energia química usada no metabolismo, dirigindo quase todos os processos celulares. Os nucleotídeos são mediadores fisiológicos em grande número de processos metabólicos. O AMPc e GMPc são reguladores de um grande número de eventos celulares. A adenosina é importante regulador do fluxo sanguíneo e atividade muscular lisa. Por fim, o nucleotídeo guanina está envolvido nos sinais de tradução e estrutura do RNA (Rudolph, 1994).
Os nucleotídeos são sintetizados endogenamente, portanto, não são classificados como nutrientes essenciais. Por outro lado, a suplementação de nucleotídeos na dieta pode proporcionar efeitos benéficos sobre o sistema imune, crescimento e desenvolvimento do intestino delgado, metabolismo de lipídeos e funções hepáticas. Assim, o termo condicionalmente essencial tem sido usado para descrever o efeito dos nucleotídeos na nutrição humana e animal. Sob estas condições, o consumo exógeno de nucleotídeos na dieta poupa o organismo a lançar mão dos custos da síntese de novo e da via de salvação (Carver & Walker, 1995).
O processo de absorção de nucleosídeos e bases nitrogenadas no enterócito ocorrem por transporte ativo (sódio dependente) ou difusão facilitada. Em animais adultos saudáveis recebendo purinas, observa-se que 90% dos nucleotídeos ingeridos são absorvidos pelo intestino (Jarvis, 1989; Savaiano e Clifford, 1978; Saviano et al., 1980; Sonoda e Tatibana, 1978).
Uauy et al. (1994) observaram que as purinas da dieta não foram significativamente incorporadas dentro do ácido nucléico hepático, mas as pirimidinas o foram. Ambos são absorvidos pelas células intestinais, mas o excesso de purina é convertido em ácido úrico.
O fígado é o maior local de síntese das purinas no organismo dos animais (Mateo, 2005). O citosol da célula contém todas as enzimas que sintetizam e degradam as purinas (Carver e Walker, 1995). Para a síntese de IMP ocorre a necessidade de 4 moléculas de ATP e 10 reações químicas, utilizando inicialmente o 5-fosforibosil 1-pirofosfato (PRPP), duas moléculas de glutamina, uma de glicina, duas moléculas de formil-tetraidrofolado e CO2 (Champe e Harvey, 1997 e Rudolph, 1994). Por fim, a conversão do IMP em AMP ou GMP utiliza uma rota de duas etapas, requerendo uma molécula de energia, ou seja, para a síntese de AMP é necessário um GTP e para a síntese de GMP é necessário um ATP.
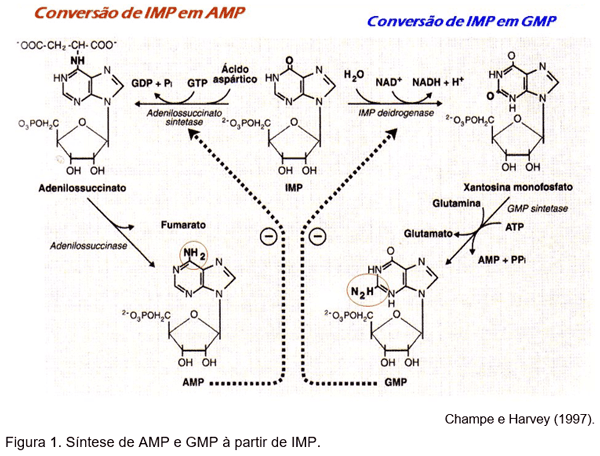
O IMP é denominado de nucleotídeo mãe (Champe e Harvey, 1997), pois serve como ponto de ramificação para a síntese de AMP e GMP (Mateo, 2005) que são aceptores de grupos fosfatos para formar ATP e GTP, os intermediários na escala bioenergética dos compostos fosfatados. (Figura 1).
A taxa de produção do AMP e GMP proporciona um feedback negativo, inibindo as enzimas adenilsuccinato sintetase e a IMP desidrogenase. A reação catalisada pela glutamina:fosforribolsil pirofosfato amidinotransferase (PRPP amidinotranferase) é também inibida pelo ATP, AMP, GTP e GMP (Mateo ,2005). Se ambos, AMP e GMP, estão presentes em quantidades adequadas a síntese de novo das purinas é desativada na etapa catalisada pela aminotransferase (Champe e Harvey, 1997).
As purinas que resultam do metabolismo normal dos ácidos nucléicos celulares, ou que são obtidas da dieta, quando não degradadas, podem ser convertidos em nucleosídeos trifosfatos e usado pelo corpo (Champe e Harvey, 1997).
Diante dos diferentes papeis fisiológicos dos nucleotídeos no organismo dos animais, Mateo (2005) e Rudolph (1994) afirmaram que o ATP é a mais abundante fonte de energia química usada no metabolismo, controlando a maioria dos processos celulares e servindo como estoque de energia (Barness, 1994). Enquanto o nível de desoxiribonucleotídeos está entre 2 a 60 µmol/L no organismo dos animais, o nível de ribonucleotídeos é maior, baseado na concentração do ATP em 2 a 10 mmol/L. Por sua vez, a concentração de outros ribonucleotídeos está entre 0,05 a 2 mmol/ L. O nível de difosfatos e monofosfatos são tipicamente menores do que os trifosfatos (Rudolph, 1994)
O transporte ativo de moléculas e íons, a síntese de macromoléculas e o trabalho mecânico são inteiramente dependentes do suprimento de energia na forma de ATP. A adenina é um importante regulador da fluidez sanguínea e atividade muscular. O AMPc e GMPc também são mediadores de numerosos processos metabólicos, regulando grande número de reações metabólicas.
Apesar dos relatos demonstrarem o potencial das purinas como fonte de energia metabólica para os animais, atualmente o IMP e o GMP são classificados como realçadores de sabor, tendo importante papel na indústria alimentícia, contribuindo no paladar umami (Mateo, 2005; Loliger, 2000 e Santos 2015), ou seja, um estímulo gustativo ocasionado por aminoácidos e nucleotídeos que interagem com as células gustativas da língua (Niki et al. 2010). Para Santos (2015) o nervo corda do tímpano que inerva a parte anterior da língua evidencia a acentuada percepção do sabor dos aminoácidos (umami, adocicado ou amargo). No paladar umami, há receptores na língua que são sensibilizados com a presença do glutamato e dos nucleotídeos inosinato e guanilato (Baldion e Flores, 2011). O IMP intensifica o sabor dos alimentos quando associado ao glutamato. Dentro da mesma descrição de efeito do nucleotídeo IMP, o AMP, também tem sido estudado no papel regulatório da inflamação e imunidade (Lapa, 2012).
Diante da necessidade de investigar o efeito dos nucleotídeos purinas como fonte de energia metabólica, relatos na nutrição esportiva humana tem classificado, por exemplo, o IMP como um importante suplemento alimentar para atletas de alto rendimento. Segundo Monteiro (2015), o IMP foi isolado no músculo pelo cientista alemão Von Justus Lieberg no século XIX. Até 1970 acreditava-se que era apenas um elemento resultante do metabolismo celular, mas médicos japoneses identificaram e aprovaram o seu valor terapêutico em pacientes com problemas cardiovasculares. Por fim, os soviéticos iniciaram o uso do IMP no mundo do atletismo, devido a sua capacidade de aumentar a energia disponível, ajudando o atleta a intensificar as repetições em treinamentos prolongados. Atualmente o IMP é considerado um dos suplementos esportivos mais importantes para atletas de alto desempenho.
O IMP também estimula a produção da 2,3 difosfoglicerato, que é essencial para o transporte de oxigênio no sangue para as células musculares na obtenção de energia, melhorando a capacidade aeróbica e anaeróbica do atleta, portanto, atua prevenindo a fadiga e ajuda na recuperação muscular (Monteiro, 2015).
Para utilizar o IMP como regenerador de energia na alimentação de frangos de corte, faz se necessário que este aditivo apresente comportamento similar às demais fontes de energia, ou seja, deve estabelecer o equilíbrio da relação nutriente:caloria da dieta sem comprometer o suprimento de ácidos graxos essenciais, a absorção de vitaminas lipossolúveis, perfil hormonal, por fim, não deve alterar, negativamente, os parâmetros de desempenho, rendimento de carcaça e de qualidade da carne das aves.
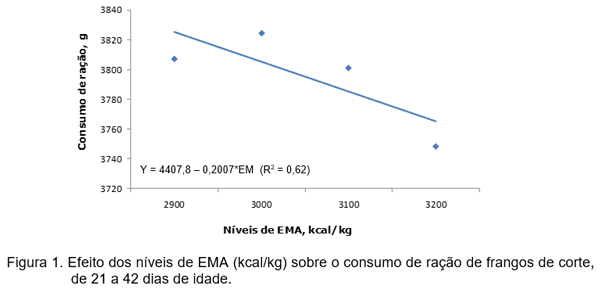
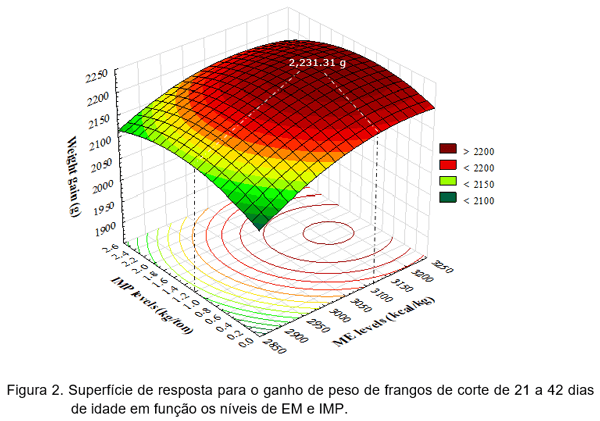
Com o objetivo de verificar o efeito interativo entre níveis de energia metabolizável (EMA) e inosina monofosfato (IMP) na alimentação de frangos de corte, Rodrigueiro et al. (2018a) utilizaram 1.200 frangos de corte machos de 21 a 42 dias de idade num delineamento inteiramente casualizado e esquema fatorial 4 x 4, ou seja, quatro níveis de EMA (2.900; 3.000; 3.100 e 3.200 kcal EMA/kg de ração) e quatro níveis de IMP (0,0; 0,8; 1,6 e 2,4 kg de IMP/t. de ração), com 5 repetições de 15 aves cada. As rações fareladas foram formuladas para atender as recomendações de Rostagno et al. (2011), exceto para EMA. Os níveis de IMP foram administrados em substituição ao inerte das dietas. As rações experimentais e a água foram fornecidas à vontade. As aves e as sobras de rações foram devidamente pesadas aos 42 dias de idade para avaliar o consumo de ração (CR), ganho de peso (GP) e a conversão alimentar (CA). As variáveis foram submetidas ao procedimento GLM do software SAS (SAS Institute, 2009) A metodologia superfície de resposta foi utilizada para estabelecer a relação entre os tratamentos estudados assim como o modelo de regressão polinomial, considerando os efeitos lineares, quadráticos e as interações dos níveis de EMA e IMP. Os autores observaram que o consumo de ração (CR) reduziu linearmente (Figura 2) com o aumento da EMA (Y=4407,8–0,2007*EMA; R²=0,62), porém não foi observado efeito significativo do IMP sobre a mesma variável.
A EMA e o IMP foram dependentes no modelo superfície de resposta (P=0,003) para o ganho de peso (GP) conforme a equação: Y=-6911,258+5,698*EMA+199,706*IMP–0,000891*EMA²–23,68952*IMP²-0,04527*IMP*EMA; R²=0,70, estimando o melhor ganho de peso em 2.231,31 gramas aos 42 dias de idade com o nível de 3.167,58 kcal de EMA/kg e 1,188 kg de IMP/t. de ração (Figura 2).
Os resultados estão de acordo com Mendes et al. (2004) os quais relatam que o nível de EMA da ração interfere no desempenho dos frangos de corte, porém, adicionalmente, esta pesquisa, estabelece que a presença do nucleotídeo IMP pode ter efeito complementar às demais fontes de energia da ração, melhorando o desempenho dos frangos de corte.
A fosforilação oxidativa das moléculas ricas em energia (carboidratos e ácido graxos) presente nas rações dos animais, são metabolizados por uma série de reações de oxidação, produzindo CO2 e H2O. Os intermediários destas reações doam elétrons para as coenzimas nicotinamida adenina dinucleotídeo (NAD+) e flavina adenina dinucleotídeo (FAD), para formar coenzimas reduzidas ricas em energia, NADH e FADH2, no ciclo do ácido cítrico. Estas coenzimas reduzidas doam um par de elétrons a um conjunto especializado de transportadores de elétrons na mitocondria. À medida que os elétrons atravessam a sua cadeia de transporte, perdem energia, e, parte desta energia é capturada e armazenada na forma de ATP, a partir de ADP + fosfato inorgânico (Champe e Harvey, 1996 e Fernandes e Toro-Velasques, 2014). O ADP, por sua vez, é um nucleotídeo aceptor de grupos fosfatos sintetizado a partir de reações de ramificação do nucleotídeo IMP (Champe e Harvey, 1996).
Diante deste exposto, é possível concluir que a suplementação exógena do nucleotídeo IMP para frangos de corte, pode, em parte, atender ou tornar mais eficiente a energia metabólica proveniente da via glicolítica, ciclo do ácido cítrico e da cadeia de transporte de elétrons, colaborando para o atendimento da exigência energética dos frangos de corte, comprovando o princípio da complementaridade entre as fontes de energia: carboidratos, gorduras e proteínas, agora, o IMP.
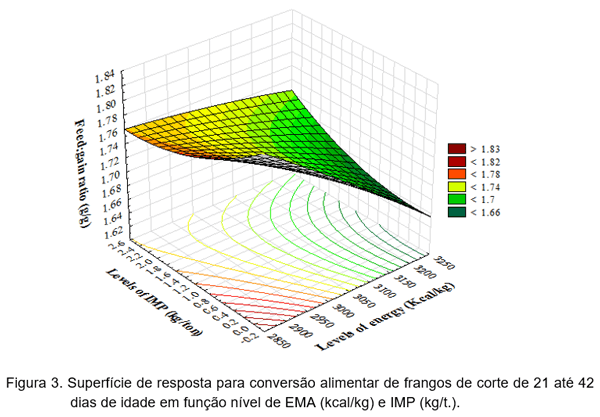
Foi observado ajuste na superfície de resposta para a CA (Y = 2,877276951–0,000369929*EMA-0,321952268*IMP+0,015854773*IMP²+0,000091653*EMA*IMP;R²=0,70). O modelo não demonstrou um ponto estacionário gerando, portanto, diferentes respostas para obter a mesma CA (Figura 3).
Para encontrar os níveis ótimos, foi realizado o desdobramento da interação de EMA vs IMP para a CA. Os dados se ajustaram às funções quadráticas para as doses de 0,0 kg até 2,4 kg de IMP/t. dentro dos níveis de EMA. As equações demonstram melhora na CA com 1,64 kg de IMP/t. e 2.900 kcal de EMA/kg (Y=1,7993-0,0581*IMP+0,0178*IMP²; R²=0,95); 1,54 kg de IMP/t. e 3.000 kcal de EMA/kg (Y=1,7837-0,0655*IMP+0,0213*IMP²; R² =0,82); 1,02 kg de IMP/t. e 3.100 kcal de EMA/kg (Y = 1,719-0,0242*IMP+0,0118*IMP²; R²=0,96); e 0,84 kg de IMP/t. para 3.200 kcal de EMA/kg (Y=1,6927-0,0204*IMP+0,0122*IMP²; R²=0,99).
Nesta pesquisa os autores concluíram que houve interação entre os níveis de IMP e a EMA sobre o ganho de peso e a conversão alimentar para frangos de corte, indicando que o IMP pode ser um aditivo complementar às fontes de energia comumente utilizadas nas dietas das aves.
Com o objetivo de validar o nucleotídeo inosina monofosfato (IMP) como aditivo complementar às fontes de energia na alimentação de frangos de corte, Rodrigueiro et al. (2018b), utilizaram 144 frangos de corte machos de 18 até 42 dias de idade em gaiolas de metabolismo, num DIC, com 6 tratamentos (2 dietas controles + 4 níveis de inosina monofosfato (IMP)) e 8 repetições de 3 aves cada. A dieta controle positivo a base de milho e farelo de soja contendo 3.125 kcal EMA/kg, foi formulada conforme Rostagno et al. (2011) para a fase de crescimento. A dieta controle negativo foi formulada com redução de 100 kcal de EMA/kg quando comparada à dieta controle positivo (3.025 kcal de EMA/kg). Os 4 níveis de IMP (0,50; 1,0; 1,5 e 2,0 kg de IMP/t. de ração) substituíram o inerte da dieta controle negativo. As aves e as sobra da ração foram pesadas aos 42 dias de idade para avaliação do consumo de ração (CR), ganho de peso (GP) e conversão alimentar (CA). As variáveis foram submetidas ao procedimento GLM do software SAS, sendo as comparações das médias realizadas pelo teste SNK (5%). Foram realizadas análises de contrastes para comparação dos tratamentos. Também, os dados obtidos de cada parâmetro foram desdobrados em polinômios ortogonais de forma a permitir a análise de regressão de acordo com suas distribuições, sem considerar o tratamento controle positivo. O melhor nível de IMP foi estabelecido pelo primeiro ponto de interseção da equação quadrática com o platô do LRP conforme Sakomura e Rostagno (2007).
O desempenho dos frangos de corte no período de 18 a 42 dias de idade submetidos aos tratamentos pode ser observado na Tabela 1, Figura 3 e 4.
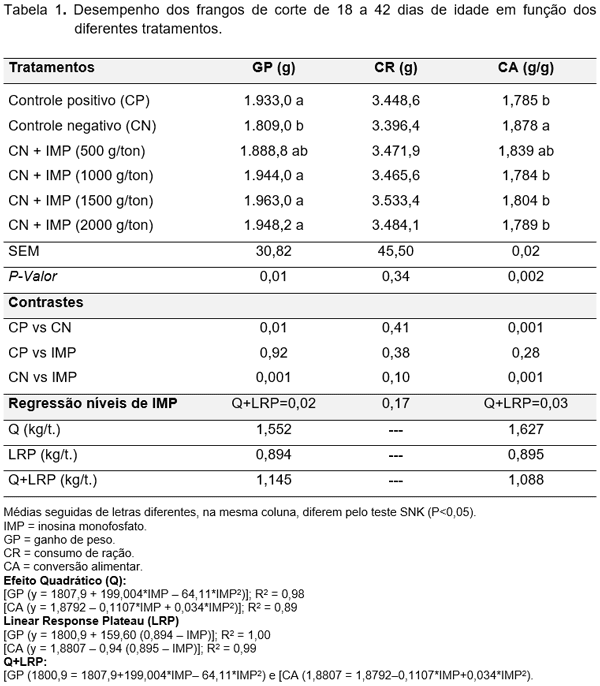
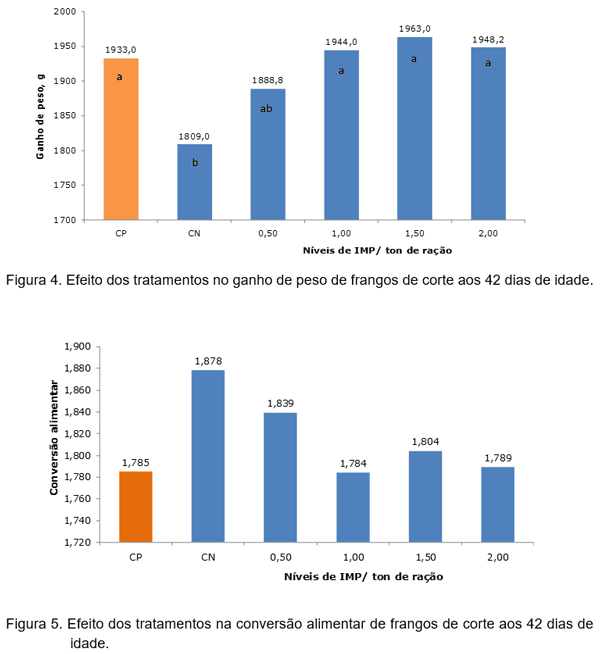
No contraste, as aves ao receberem a ração com 3.025 kcal de EMA/kg reduziram o GP e piorou a CA sem afetar o CR. O IMP melhorou o GP (P=0,001) e a CA (P=0,001) de frangos quando comparado com aqueles alimentados com 3.025 kcal de EMA/kg. O IMP na dieta de 3.025 kcal de EMA/kg melhorou o desempenho de forma a atingir o mesmo GP e a mesma CA de aves alimentadas com 3.125 kcal de EMA/kg (P≤0,05), justificando a contribuição energética ao redor de 100 kcal de EMA/kg de ração. Os tratamentos não afetaram o CR (P≥0,05). A recomendação dada pelo ponto de inserção da equação quadrática com o LRP foi de 1,145 kg IMP/t. e 1,088 kg IMP/t. para obter o melhor GP e a melhor CA, respectivamente. O resultado demonstrou que o IMP pode ser um importante elemento que, ao ser adicionado na dieta, suprirá pelo menos em parte as necessidades energéticas diárias das aves. Para Mateo (2005) o IMP é um intermediário do metabolismo energético dos animais e serve como ponto de ramificação para a síntese de AMP e GMP, os aceptores de grupos fosfatos para formar ATP e GTP (intermediários na escala bioenergética dos compostos fosfatados). Para Leeson & Summers (2005) o ATP é formado pela união de uma molécula de ácido fosfórico com uma molécula de adenosina difosfato (ADP). O aprisionamento da energia pelo ADP no metabolismo da glicose e ácido graxo não somente reduz a perda de calor (incremento calórico), mas fornece um constante suprimento de energia corporal. O resultado deste experimento justifica a contribuição energética do IMP de 100 kcal de EMA/kg de ração.
Conclusivamente o IMP não dever ser classificado unicamente como um componente que realça o sabor dos alimentos, enaltecendo o paladar umami (Mateo, 2005; Loliger, 2000 e Santos 2015), mas, através deste artigo, é possível afirmar que o IMP pode ser um importante elemento que, quando adicionado na dieta, suprirá pelo menos em parte a necessidade energética diária das aves, atingindo eficiente ganho de peso e conversão alimentar. O uso do nucleotídeo inosina monofosfato na alimentação de frangos de corte abre evidências para entender o princípio da complementaridade das fontes de energia na nutrição das aves.











.jpg&w=3840&q=75)