Introdução
Na produção industrial de ovos para incubação, buscam-se práticas para concentrar a postura nos ninhos, evitar a postura na cama e, conseqüentemente, a presença de ovos sujos. Tanto o manejo dos ninhos na pré-postura como a utilização de um número adequado de coletas diárias, contribuem na obtenção de ovos limpos e com menor grau de contaminação. Além disso, o peso do ovo e a qualidade da casca (gravidade específica) são fatores importantes na produção do ovo fértil. Considera-se que maior gravidade específica resulta em melhor qualidade de casca e, conseqüentemente, em ovos mais apropriados para incubação. Salienta-se, no entanto, a importância da relação entre peso do ovo e gravidade específica, em que o peso do ovo aumenta e a gravidade específica diminui com a idade das reprodutoras.
Cave (1981) e Bootwalla et al. (1983) colocam em dúvida se o esquema de alimentação única pela manhã atende à demanda por nutrientes, especialmente no momento da formação da casca do ovo. Cave (1981), trabalhando com matrizes pesadas, constatou que a alimentação duas ou três vezes ao dia permitiu melhora no peso e na produção de ovos e maior eficiência na utilização do alimento.
Farmer & Roland Sr (1983) e Bootwalla et al. (1989) relataram que os diferentes horários de alimentação influenciaram o ciclo de formação do ovo e os níveis de cálcio e fósforo do sangue no momento da calcificação e que o horário de maior necessidade de cálcio, para a formação da casca do ovo, ocorreu no período noturno. No entanto, Brake (1988) observou que tanto a produção quanto o peso do ovo não foram influenciados pelo horário da alimentação.
De acordo com Summers (1987) e Rutz (1992), o catabolismo do excesso de proteína em ácido úrico produz mais calor corporal, ocasionando estresse adicional ao das elevadas temperaturas. Aves alimentadas às 6h apresentaram maior incremento calórico cinco horas após a refeição que aquelas alimentadas às 14h (Wilson et al., 1989). Incremento calórico promove alta ofegação e perda de CO2, podendo tornar o problema ainda mais grave para as reprodutoras em virtude da temperatura ambiental. Como recurso para perda de calor ou redução da temperatura corporal, surgem problemas na produção de ovos decorrentes da falta do CO2 na principal reação de formação do carbonato de cálcio, necessário para a calcificação da casca do ovo.
Neste contexto, realizou-se este experimento com o objetivo de associar o horário de fornecimento do alimento às aves com a produção de ovos por coleta diária, peso e gravidade específica do ovo em reprodutoras de frangos de corte.
Material e Métodos
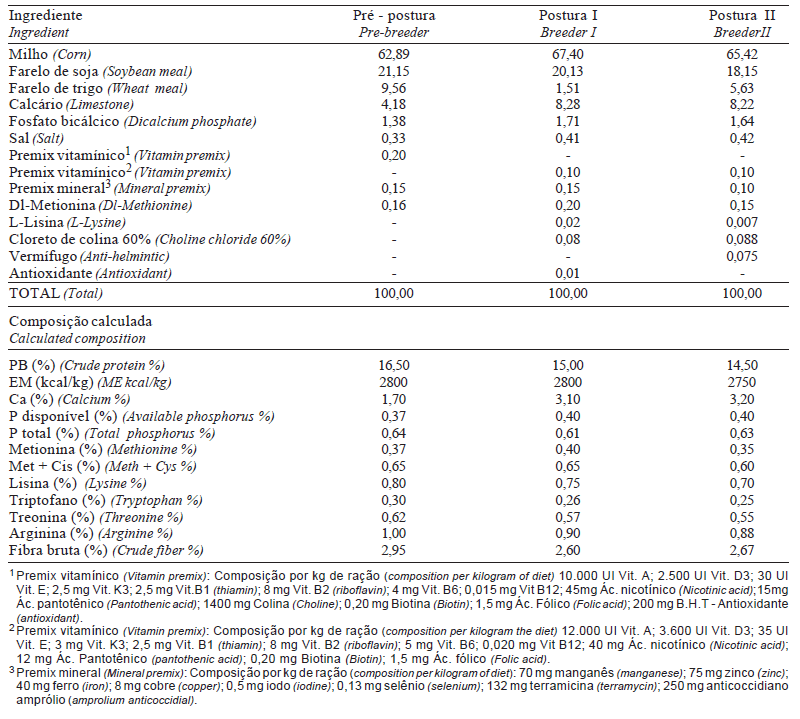
Utilizou-se um aviário experimental nas dependências da Embrapa Suínos e Aves para alojar, a partir das 20 semanas de idade, 1.296 fêmeas da linhagem Arbor Acres e 144 machos de outra linhagem comercial, distribuindo-se 27 fêmeas e três machos por boxe, no período de 24 a 66 semanas de idade. Para se evitar o estresse, a ração (peletizada e posteriormente moída) a ser fornecida no dia seguinte era pesada e os comedouros levantados logo após a refeição do dia. Para as fêmeas, a quantidade de ração diária foi baseada no peso corporal e na produção de ovos, e para os machos, tomou-se como base apenas o peso corporal. O peso corporal foi obtido em intervalos de duas semanas, utilizando-se a média de quatro boxes por tratamento. Os tratamentos consistiram de quatro horários de fornecimento do alimento (arraçoamento único, às 6h30; arraçoamento dual (50% às 6h30 e 50% às 15h30); arraçoamento único, às 11h; e arraçoamento único, às 15h30), iniciados a partir da 20ª semana, para adaptação das aves. Para se evitar cama úmida, houve restrição de água durante o período experimental, que foi fornecida de 6h30 às 9h, de 11h às 13h30 e de 15h30 às 18h. Eventualmente, em dias quentes, não houve restrição. Foram adotados os seguintes períodos para cada tipo de ração: pré-postura, de 18 a 23 semanas, postura I, de 24 a 47 semanas e postura II, de 48 a 66 semanas de idade. A debicagem, o programa de luz e as exigências nutricionais atenderam as recomendações do manual da linhagem Arbor Acres (Sadia, 1993). A porcentagem média dos ingredientes das dietas experimentais das reprodutoras encontram-se na Tabela 1.
Neste trabalho, avaliou-se a classificação dos ovos produzidos em seis coletas diárias, realizadas às 6h30, 9h, 11h, 13h30, 15h30 e 17h30.
Em cada boxe, a porcentagem total de ovos (PTOTAL) por coleta foi obtida pelo quociente entre o número total de ovos da coleta da semana e o somatório das reprodutoras sobreviventes em cada dia durante a semana. A porcentagem total de ovos foi calculada pela média das produções durante o período de 25 a 66 semanas. O cálculo dos percentuais de produção para ovos no ninho (PNINHO), limpos (PLIMPO) e quebrados (PQUEMO) obedeceu aos mesmos critérios da variável PTOTAL. Os ovos quebrados e membranosos foram agrupados na variável PQUEMO. Como a porcentagem de ovos de cama é complemento dos ovos de ninho e a porcentagem de ovos sujos é complemento dos ovos limpos em relação à porcentagem total de ovos produzidos, foram apresentadas e discutidas apenas as variáveis porcentagem de ovos de ninho e porcentagem de ovos limpos.
Tabela 1 - Composição percentual e calculada da dieta
Considerou-se como variáveis caracterizadoras da qualidade do ovo incubável o peso e a gravidade específica.Para avaliar o peso médio do ovo (PESOVO) e a gravidade específica (GE), foram utilizados todos os ovos produzidos em cada boxe, nos dois últimos dias das semanas 28, 32, 36, 40, 44, 48, 52, 56, 60 e 65. As coletas foram redefinidas para três, após terem sido reagrupadas duas a duas, obedecendo a seqüência natural inicialmente estabelecida.
Para determinação da GE, após pesagem, os ovos foram colocados dentro de uma cesta e mergulhados em 10 baldes com soluções de densidade diferentes, calibradas por um densímetro, variando a concentração de 1,062 a 1,098, com incrementos de 0,004 g/mL. Este procedimento foi realizado para cada boxe, efetuando- se, seqüencialmente, a avaliação para todas as coletas dentro do mesmo, antes de iniciar as avaliações no próximo boxe.
Na análise das variáveis PTOTAL, PNINHO, PLIMPO, PQUEMO, PESOVO e GE, adotou-se o procedimento GLM do SAS (1996) envolvendo o seguinte modelo para parcelas sub-subdividida:
yijlk = μ + ti + eij + cl + ctil + eijl + sk + tsik + cslk + tcsilk + eijlk
em que i = 1,2,3,4 tratamentos; j = 1,...,12 boxe;l = 1,2,...,C coletas diárias; k = 1,..., K semanas de avaliação, de modo que: yijlk é o valor da resposta pertencente ao boxe ij na coleta l da semana k; μ, a média da resposta no experimento; ti, o efeito do tratamento i; eij, o erro experimental suposto seguir a distribuição normal de média 0 e variância constante s2 ; cl , o efeito de coleta; ctil, o efeito da interação tratamento i x coleta l; eijl, o erro de avaliação decorrente da coleta, suposto seguir a distribuição normal de média zero e variância constante σc 2 ;sk, o efeito de semana k; tsik, o efeito da interação tratamento i x semana k; cslk, o efeito de interação coleta l x semana k; tcsilk, o efeito de interação tratamento i x coleta l x semana k; e eijlk, o erro de avaliação semanal suposto seguir a distribuição normal de média zero e variância constante σs 2. O erro experimental eij é o termo usado para testar hipóteses sobre o efeito de tratamentos; eijl, o termo usado para testar hipóteses sobre coletas e interação tratamentos x coletas; eijlk, o termo usado para testar hipóteses sobre efeito de semanas e as interações tratamentos x semanas, coletas x semanas e tratamentos x coletas x semanas.
As médias dos tratamentos e das coletas foram comparadas pelo teste T de Student, enquanto, para a idade das reprodutoras, utilizou-se análise de regressão polinomial.
A inclusão completa das interações duplas e tripla entre os fatores tratamentos, semanas e coletas, no modelo usado, teve como finalidade principal purificar o erro experimental, para aumentar a eficiência do experimento.
Como a interação coletas x semanas e a interação tripla tratamentos x coletas x semanas são desprovidas de qualquer interesse prático, ambas não foram desdobradas. Deu-se ênfase aos efeitos principais dos fatores envolvidos e às interações de tratamentos com coletas e de tratamentos com semanas.
Resultados e Discussão
Houve efeito (p≤0,05) para as variáveis PTOTAL, PNINHO e PLIMPO em relação aos fatores tratamentos, coletas, semanas e às interações entre os fatores.
Para a variável PQUEMO, não foi evidenciado efeito de tratamentos e de interação tratamentos x semanas. Este resultado contraria os relatos de Robinson et al. (1995), que constataram alta incidência de ovos com casca deformada quando trabalharam com aves alimentadas à vontade.
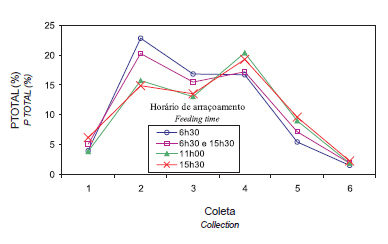
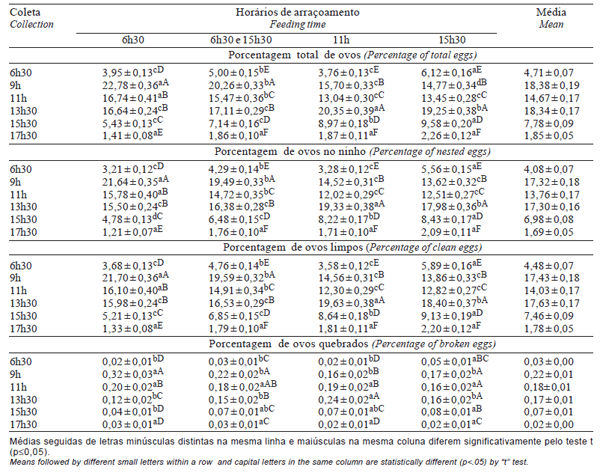
As interações tratamentos e coletas para PTOTAL, PNINHO e PLIMPO foram semelhantes entre si e seus percentuais médios por coleta encontram- se na Tabela 2. Consta na Figura 1 ilustração gráfica considerando tratamento e coleta para PTOTAL; as demais variáveis apresentaram comportamento semelhante.
Com exceção de ovos quebrados, nas coletas das 9h e 11h, as aves com arraçoamento único às 6h30 apresentaram comportamento diferente e superior (p≤0,05) àquelas dos outros tratamentos. Verificouse que os percentuais de produção de ovos dos demais tratamentos foram deslocados para as coletas mais tardias em virtude da interferência do horário de arraçoamento.
As aves que receberam todo o alimento às 6h30, ou parte às 6h30 (dual) tiveram o maior percentual de produção de ovos coletado às 9h, enquanto aquelas que receberam todo o alimento às 11h e às 15h30 apresentaram o maior percentual de produção de ovos coletados às 13h30, demonstrando que o horário de arraçoamento influencia no horário de postura das aves das aves (Figura 1).
Figura 1 - Perfil do percentual de produção total de ovos (PTOTAL) referente a seis coletas em reprodutoras de frangos de corte, de acordo com o horário de arraçoamento.
Tabela 2 - Percentuais médios de produção total de ovos, ovos de ninho, ovos limpos e ovos quebrados nos diferentes horários de arraçoamento e de coleta em reprodutoras de frangos de corte
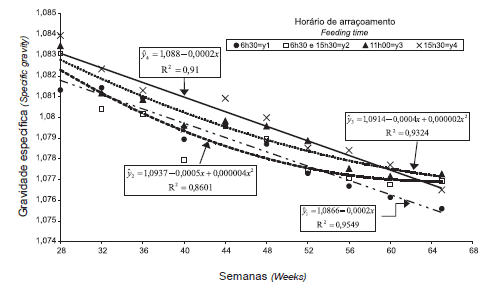
Figura 2 -Perfil da gravidade específica do ovo de acordo com o horário de arraçoamento.
A produção de ovos registrada na primeira coleta é influenciada pelo grupo de aves que realizaram a postura após a última coleta do dia anterior, ou seja, é maior para aves com arraçoamento às 15h30 e dual, possivelmente por terem recebido pelo menos parte do alimento mais tarde, o que está de acordo com Samara et al. (1996), que constataram produção de 59,2% dos ovos antes das 14h para as reprodutoras alimentadas às 7h e de 53% para aquelas alimentadas às 18h, caracterizando atraso na postura desse grupo de aves.
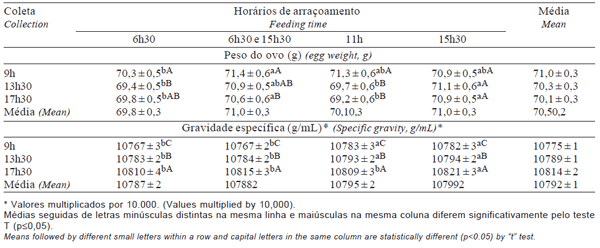
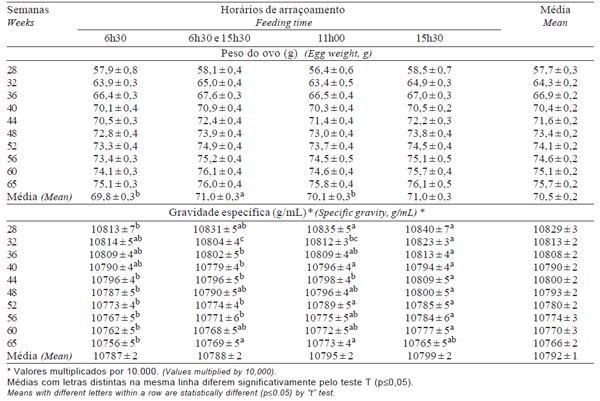
As médias para tratamento dentro de coleta e para tratamento dentro de semana, para PESOVO e GE podem ser observadas nas Tabelas 3 e 4, respectivamente. Houve interações dupla e tripla (p≤0,05) entre tratamentos, coletas e semanas, exceto para tratamentos vs semanas, em relação ao PESOVO, e para tratamentos vs coletas vs semanas, em relação a GE.
Tabela 3 - Médias do peso do ovo e da gravidade específica por coleta nos diferentes horários de arraçoamento
Tabela 4 - Médias para peso e gravidade específica do ovo por semana para as reprodutoras nos diferentes horários de arraçoamento
Para PESOVO, os tratamentos ocasionaram comportamento diferente em cada coleta, entretanto, as aves que receberam o arraçoamento dual e aquelas que receberam o arraçoamento único às 15h30 apresentaram, em qualquer coleta, o PESOVO no mínimo igual aos demais, justificando a maior média geral desses tratamentos. Resultados semelhantes foram constatados por Bootwalla et al. (1983), Farmer & Roland Sr (1983), Yannakopoulos & Tserveni-Gousi (1987) e Brake (1988). Este aumento no PESOVO pode ter decorrido da maior eficiência na utilização do alimento. Por sua vez, as aves dos dois tratamentos que receberam todo o alimento pela manhã não foram favorecidas com a mesma intensidade, possivelmente em razão da conhecida correlação negativa entre produção e PESOVO, o que pode ter afetado as aves alimentadas às 6h30. O estresse pelo calor, resultante das temperaturas iguais ou superiores a 30oC na maioria das semanas, e o calor metabólico podem ter exercido maior influência sobre as aves alimentadas às 11h. Entretanto, Harms (1991b) não identificou diferença (p≤0,05) no PESOVO quando alimentou matrizes de frangos de corte às 8h e ou 16h no período de 52 a 62 semanas de idade.
O PESOVO decresceu entre a coleta de 9h e a de 17h30, indicando que ovos coletados nas primeiras horas do dia tendem a ser mais pesados, confirmando os relatos de Brake (1985), que constatou que o PESOVO foi maior quando a postura ocorreu antes das 9h. Por outro lado, não coincide com os resultados de Yannakopoulos & Serveni-Gousi (1987), que verificaram menor peso para ovos postos e coletados pela manhã.
Independentemente dos tratamentos, o PESOVO tende a aumentar com a idade das aves, seguindo a equação quadrática: Yˆ = 15,741 + 2,0355x - 0,0173x2 (R2 = 0,978), sugerindo que galinhas mais velhas produzem ovos mais pesados (Yannakopoulos & Serveni-Gousi, 1987).
Para GE, as interações tratamentos x coletas e tratamentos x semanas fizeram com que o efeito não fosse o mesmo para as fontes de variação. Entretanto, em razão do efeito altamente significativo para tratamentos, a GE aumentou com o horário da coleta e foi, na maioria das vezes, maior para as aves arraçoadas às 11h e às 15h30, que para as demais, independentemente de coletas e semanas (Tabela 3).
A GE diminuiu com a idade das aves (Tabela 4 e Figura 2), de forma similar ao observado por Yannakopoulos & Serveni-Gousi (1987), que verificaram que ovos coletados à tarde apresentaram maior gravidade específica que aqueles coletados pela manhã e que galinhas mais velhas produzem ovos com GE menor e, conseqüentemente, com casca mais fina.
Farmer & Roland Sr (1983), observando os níveis de Ca nos diferentes segmentos do aparelho digestivo, constataram que reprodutoras alimentadas à tarde, tiveram mais Ca disponível durante o estágio de calcificação do ovo que as reprodutoras alimentadas pela manhã. Neste trabalho, parece que a disponibilização do Ca para a glândula calcífera foi menor nas aves arraçoadas às 6h30 e dual no momento da formação da casca, o que confirma os estudos de Brake (1985), que constatou que a GE foi menor para ovos postos e coletados de 9h às 13h e maior para ovos postos e coletados de 15h às 19h, razão pela qual o autor sugere a existência de variação na GE dos ovos postos no decorrer do dia, em função do acesso ao alimento.
A GE foi beneficiada (p≤0,05) com o arraçoamento mais tardio. Resultados semelhantes foram constatados por Farmer & Roland Sr (1983), Bootwalla et al. (1983), Brake (1985), Yannakopoulos & Tserveni- Gousi (1987), Brake (1988) e Harms (1991b). A menor GE para ovos de reprodutoras postos e coletados de 9h e 13h, associada ao menor PESOVO, observado por Brake (1985), se assemelha aos resultados médios obtidos neste trabalho, em que se constatou aumento na GE nos ovos postos e coletados a partir de 9h, indicando que ovos coletados nas primeiras horas do dia apresentam cascas mais finas. Estes dados confirmam os de Roland Sr & Harms (1974), que verificaram que a GE e o PESOVO estão relacionados ao horário de postura e que a melhoria na qualidade da casca dos ovos postos à tarde não depende da redução do peso dos mesmos.
Verifica-se que geralmente ovos mais pesados indicam menor GE. No entanto, os dados revelam que igual PESOVO necessariamente não está associado à igual GE. O inverso é verdadeiro, pois semelhantes GE não necessariamente apresentaram o mesmo PESOVO, corroborando as observações de Roland Sr & Harms (1974).
Considerando as médias de PESOVO e GE das aves que receberam arraçoamento dual, estes dados são similares aos obtidos por Brake (1988), que observou que o arraçoamento mais tarde do dia apresentou considerável aumento no PESOVO com redução na GE, fato atribuído ao aumento do peso da gema e do albúmen. Portanto, outro fator além de PESOVO passou a influenciar na qualidade da casca, que possivelmente tenha sido o efeito do horário de arraçoamento.
As aves com arraçoamento dual, embora tenham recebido 50% da alimentação à tarde, não responderam à GE do mesmo modo que as aves arraçoadas às 11h e às 15h30. Porém, esse tratamento favoreceu o aumento do PESOVO, possivelmente por disponibilizar nutrientes para maior síntese de gema e albúmen, o que confirma os estudos de Cave (1981), que verificou vantagem no esquema de alimentação mais vezes ao dia, em razão do aumento na produção de ovos mais pesados durante o período crítico, nas primeiras semanas de postura, e maior média de PESOVO para o período geral. Os resultados de Samara et al. (1996) também foram semelhantes, pois confirmaram que a mudança do horário de alimentação para a tarde não foi efetiva nas aves submetidas ao estresse de calor. No entanto, a alimentação em duas refeições (50% da ração às 7h e 50% às 18h) foi mais eficiente, como no presente trabalho.
O peso do ovo aumentou com a idade das reprodutoras, como descrito por O´Sullivan et al. (1991), que verificaram a mesma resposta para peso do povo. Os autores também constataram aumentos para peso do albúmen, da gema e da casca, de modo que o elevado peso do albúmen foi atribuído ao incremento no conteúdo de água e da casca e gema, ao acúmulo de matéria seca.
Conclusões
Os sistemas de arraçoamento mais tardios provocaram atraso e diluição da postura das reprodutoras ao longo do dia. O sistema de arraçoamento dual pode ser uma alternativa ao sistema de arraçoamento único praticado nas primeiras horas do dia.
Literatura Citada
BOOTWALLA, S.M.; WILSON, H.R.; HARMS, R.H. Performance of broiler breeders on different feeding systems. Poultry Science, v.62, n.12, p.2321-2325, 1983.
BOOTWALLA, S.M.; WILSON, H.R.; HARMS, R.H. Plasma calcium and phosphorus levels of broiler breeders with different feeding schedules. Poultry Science, v.39, n.2, p.391-398, 1989.
BRAKE, J. Relationship of egg weight, specific gravity, and shell weight to time of oviposition and feeding in broiler breeders. Poultry Science, v.64, n.11, p.2037-2040, 1985.
BRAKE, J. Relationship of time and strain to egg shell quality and hatchability in broiler breeders. Poultry Science, v.67, n.4, p.538-543, 1988.
CAVE, N.A. Effect of diurnal programs of nutrient intake on performance of broiler breeder hens. Poultry Science, v.60, n.6, p.1287-1292, 1981.
FARMER, M.; ROLAND SR, D.A. Calcium metabolism in broiler breeder hens. 2.The influence of the time of feeding on calcium status of the digestive system and eggshell quality in broiler breeders. Poultry Science, v.62, n.3, p.465-471, 1983.
HARMS, R.H. The influence of changing time of feeding on performance of broiler breeder hens. Poultry Science, v.70, n.8, p.1695-1698, 1991b.
O´SULLIVAN, N.P.; DUNNINGTON, E.A.; SIEGEL, P.B. Relationships among age of dam, egg components, embryo lipid transfer, and hatchability of broiler breeder eggs. Poultry Science, v.70, n.10, p.2180-2185, 1991.
ROBINSON, F.E.; ROBINSON, N.A.; HARDIN, R.T. The effects of 20-week body weight and feed allocation during early lay on female broiler breeders. Journal of Applied Poultry Research, v.4, n.2, p.203-210, 1995.
ROLAND SR, D.A.; HARMS, R.H. Specific gravity of eggs in relation to egg weight and time of oviposition. Poultry Science, v.53, n.4, p.1494-1498, 1974.
RUTZ, F. Efeito da alimentação e nutrição da matriz sobre a qualidade dos pintos. In: MINI-SIMPÓSIO COLÉGIO BRASILEIRO DE NUTRIÇÃO ANIMAL, 7., 1992, Campinas. Anais... Campinas: Colégio Brasileiro de Nutrição Animal, 1992. p.39-56.
SADIA. Agropastoril Catarinense Ltda. Manual Arbor Acres. Faxinal dos Guedes, SC: Sadia, 1993. 26p.
SAMARA, M.H.; ROBBINS, K.R; SMITH, M.O. Interaction of feeding time and temperature and their relationship to performance of the broiler breeder hen. Poultry Science, 75, n.1, p.34-41, 1996.
STATISTICAL ANALYSES SYSTEM - SAS. System for microsoft Windows. release 6.12, Cary:1996. CD-ROM.
SUMMERS, J.D. Feeding and management of broiler breeders. In: CONGRESSO LATINOAMERICANO DE AVICULTURA, 10., 1987, Buenos Aires. Proceedings... Buenos Aires: ALA/ CAPIA, 1987, p.29-37.
WILSON, H.R.; MATHER, F.B.; BRIGMON, R.L. et al. Feeding time and temperature interactions in broiler breeders. Poultry Science, v.68, n.5, p.608-616, 1989.
YANNAKOPOULOS, A.L.; TSERVENI-GOUSI, A.S. Relationship of parents´ age, hatching egg weight, and shell quality to day-old chick weight as influenced by oviposition time. Poultry Science, v.66, n.5, p.829-833, 1987.