Fitase e suas implicações na digestão e absorção de nutrientes
Publicado: 18 de novembro de 2013
Por: Gilson Alexandre Gomes, Gerente Técnico - AB Vista Feed Ingredients.
Sumário
Historicamente as fitases têm sido utilizadas com o intuito de liberação de fósforo através da hidrólise do fitato. Novos estudos têm demonstrado o efeito antinutricional do fitato, e o conceito de “nutrição na ausência do fitato” principalmente através de “Superdosagens” de fitase estão trazendo ganhos significativos em desempenho animal.
Introdução
O uso de fitases em rações para animais não ruminantes é bastante elevada, e estima-se que cerca de 90% das dietas de aves e suínos contenham a adição dessa enzima. Existem 4 principais tipos de fitase no mercado que derivam de diferentes microrganismos: Peniophora, Aspergillus, E Coli selvagem e E Coli geneticamente modificada. Cada uma delas tem características diferentes em termos de estabilidade térmica, estabilidade gástrica, afinidade ao substrato (fitato) e atuação em diferentes pH’s, e são essas diferenças que fazem com que cada tipo de fitase atue de maneira diferente, e por consequência, causem diferentes liberações de nutrientes. Contudo, é um objetivo comum de todas as fitases a quebra do fitato dos ingredientes vegetais que não é disponível para os animais, assim liberando o fósforo e reduzindo a necessidade de inclusão de fosfato inorgânico (ou outras fontes de fósforo, como por exemplo, a farinha de carne e ossos) às dietas de aves e suínos, trazendo assim um benefício econômico quando da formulação das dietas. Nos últimos anos a comunidade cientifica vem discutindo os efeitos negativos que o fitato pode causar para a digestibilidade de microminerais (Harland e Morris, 1995), proteína (Cowieson e Cowieson, 2011), aminoácidos (Cowieson e Ravindran 2007, Cowieson et al., 2004), sódio (Cowieson et al., 2004) e utilização da energia. Essa é uma área de estudo relativamente nova, onde doses mais altas de fitato têm sido empregadas para determinar o efeito extra-fosfórico das fitases, ou seja, a resposta à adição de altas doses de fitase que não estejam meramente associadas à liberação de fósforo do fitato, mas sim por evitar os danos causados pelo fitato na digestibilidade dos nutrientes supracitados.
Fitato e seus efeitos anti-nutricionais
De acordo com Santos (2012) o fitato (mio-inositol 1,2,3,4,5,6-hexa fosfato desidrogenado) além de ser uma importante fonte de fósforo para nutrição animal, é também uma fonte de reserva desse mineral nos grãos, e mais recentemente se demonstrou que atua como antioxidante nos grãos através da quelação de minerais como ferro, cálcio e zinco, prevenindo o estresse oxidativo e assim evitando a morte do embrião.
Santos (2012) ainda cita que a afinidade do fitato a cátions no trato gastrintestinal varia de acordo com o pH, fato este que está relacionado com a constante de dissociação da molécula (pKa), o que significa que o fitato pode estar levemente carregado negativamente (em pH ácido) ou fortemente carregado negativamente (em pH neutro/alcalino). Desta maneira a afinidade do fitato aos cátions da dieta muda de acordo com o pH (Figura 1).
Figura 1 – Solubilidade do fitato em pH 2,0 a 7,0 na presença de Ca em concentração similar ao encontrado no trato gastrintestinal dos animais (Fonte: Universidade de Maryland, dados não publicados)

Tamin e Angel (2003) demonstraram que a baixa digestibilidade do P fítico do fitato não é devido ao fato que as aves e suínos não possuírem uma enzima capaz de hidrolisar o fitato. De acordo com os pesquisadores isto ocorre devido ao fato que alguns minerais reagem com o fitato insolubilizando-o, e assim impossibilitando a assimilação do P fítico, assim como do mineral que está ligado ao fitato. Tamin et al. (2004) demonstraram que o desaparecimento do P fítico em dietas sem calcário foi ao redor de 69%, enquanto que com somente 0,5% de calcário houve uma diminuição para 25%. Neste mesmo estudo se demonstrou que as fitases são eficientes em incrementar a absorção não somente do fósforo, mas também do cálcio.
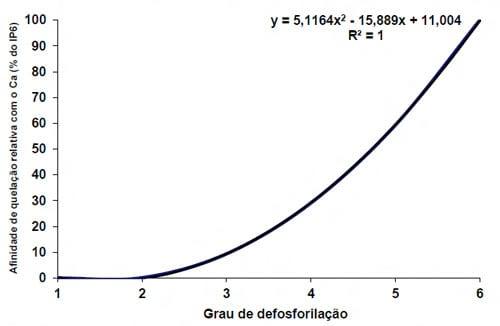
Apesar da afinidade do fitato ser maior ao cobre e zinco (Selle e Ravindran, 2007) em dietas de aves existe uma alta concentração de cálcio, e por isso a formação de complexos Ca-fitato é inevitável. Nelson (1984) demonstrou que os requerimentos de cálcio incrementam de acordo com a quantidade de fitato na dieta, e Lutrell (1993) demonstrou que a afinidade do éster IP4 (fitato com 4 grupos fosfato) ao cálcio é 70% menor que um éster IP6 (Figura 2 e 3).
Figura 2 – Requerimento de cálcio total de frangos de corte até 21d de idade alimentados com dietas purificadas e recebendo níveis crescentes de P fítico (adaptado de Nelson, 1984)
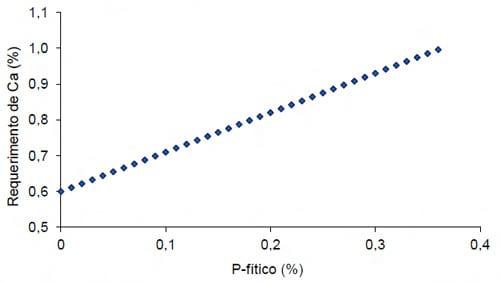
Figura 3 – Afinidade de diferentes ésteres de fitato pelo cálcio (adaptado de Lutrell, 1993)
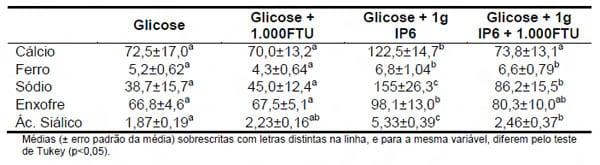
Cowieson et al. (2004) elucidaram em partes o efeito anti-nutricional do fitato quando em seus estudos perceberam que existia um incremento na produção de mucina, e na excreção de sódio e outros minerais quando frangos de corte recebiam dietas com fitato (Tabela 1). De acordo com os autores isto ocorre porque o organismo secreta mais HCl para a ativação da pepsina, já que o fitato diminui a hidrólise das proteínas e os animais tentam compensar essas perdas de digestibilidade.
Tabela 1 – Efeito do ácido fítico e fitase na excreção de minerais endógenos e de ácido siálico (mg/ave/48h) em frangos de corte (Adaptado de Cowieson et al., 2004)
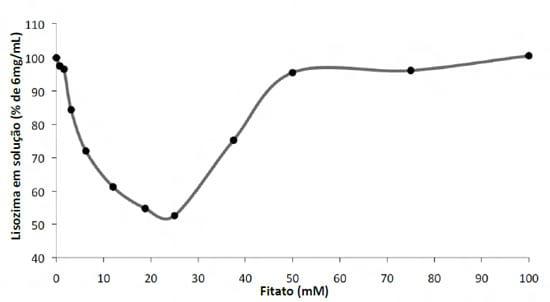
Kies et al. (2006) em ensaios in vitro demonstraram que a solubilidade de diferentes proteínas pode ser influenciada pela concentração de fitato. Neste estudo a proteína de pior solubilidade foi a do farelo de arroz, matéria prima esta que já tem um alto conteúdo de fitato em sua composição causando sua “auto-insolubilização” (Figura 4). Mais recentemente Cowieson e Cowieson (2011) propuseram que o fitato diminui a solubilidade da proteína através da atração de moléculas de água para perto de si (interação eletrostática), o que faria que os nutrientes tivessem menos água ao seu redor, e assim diminuindo a sua solubilidade/digestibilidade. (Figura 5).
Figura 4 – Impacto do pH e da concentração de fitato na solubilidade da proteína de diferentes ingredientes (adaptado de Kies et al., 2006)
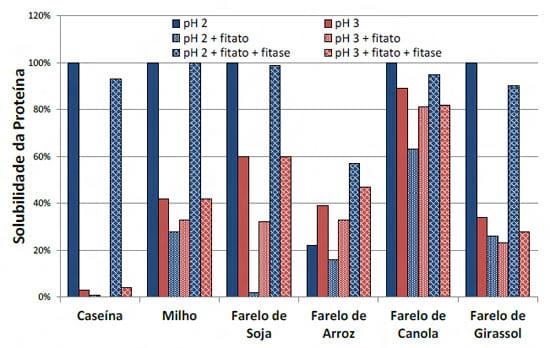
Figura 5 – Efeito da concentração crescente de ácido fítico sobre a solubilidade da lisozima (adaptado de Cowieson e Cowieson, 2011)
Análise de fitato em matérias-primas
Como mencionado acima, o fitato dos grãos desempenha um forte efeito antinutricional para as aves, e desta maneira é crucial o conhecimento da quantidade de fitato nos ingredientes e rações, e com isso tentar otimizar a utilização das fitases, empregando doses adequadas à quantidade de substrato.
Existem muitas metodologias utilizadas para a determinação da quantidade de fósforo fítico (precipitação por ferro, HPLC e metodologia enzimática), porém, o maior limitante da análise de fitato são os custos analíticos elevados. Uma alternativa para a redução dos custos seria a implementação de análise de fitato por NIRS (Near-Infrared Reflectance Spectroscopy).
Nas Tabelas Brasileiras para Aves e Suínos (Rostagno et al., 2011) podemos observar alguns valores de referencia para o conteúdo de fósforo fítico em matériasprimas brasileiras. No entanto, Wadt et al. (2010) demonstraram que a quantidade de fósforo fítico em amostras de milhos brasileiros apresentaram um valor mínimo de 0,10% e máximo de 0,26% (n=94), e concluíram que esta variação pode estar correlacionada à fertilização feita no solo, assim como por diferenças ambientais e genéticas. Em um estudo com genética de feijões (Phaseolus vulgaris) Coelho et al. (2002) verificaram que a fertilização por fósforo e a genética influenciaram a biossíntese de fitato pela planta.
Fitases
De acordo com Krabbe (2012) para que a utilização das enzimas seja bem sucedidad algumas condições são essenciais: presença do substrato, presença da enzima específica para aquele substrato, relação adequada entre atividade enzimática e quantidade de substrato, ambiente adequado para a enzima (temperatura, pH e tempo). Entretanto, muitas vezes as condições no aparato intestinal das aves não são ideais para a atividade enzimática. Onyango et al. (2005), estudando a atividade residual de diferentes fitases (bacteriana de Escherichia coli, e fúngica de Peniophora lycii) verificaram que a fitase bacteriana apresentava atividade muito maior em todos os segmentos do trato gastrintestinal de frangos de corte quando comparadas à fitase fungica. Os resultados encontrados por Onyango et al. (2005) podem ser explicados pelos resultados encontrados por Igbasan et al. (2000) que em ensaios in vitro expuseram diferentes fitases fúngicas e bacterianas a um ambiente gástrico (simulando as condições do estômago/proventrículo – pH 2 + pepsina e pancreatina) e concluíram que as E. coli fitases foram as únicas que resistiram grandemente ao desafio imposto (pH baixo e proteases endógenas).
A cinética de atuação das fitases também é outro ponto de extrema relevância. Prata et al. (2007) compararam uma fitase E. coli geneticamente modificada com outras fitases fúngicas, testando a velocidade de hidrólise do fitato e seus diferentes ésteres (IP6, IP5, IP4, IP3, IP2) em dois diferentes pH’s (2,5 e 3,5) e concluiu que a E. coli geneticamente modificada foi muito mais rápida na hidrólise do substrato, principalmente em hidrolisar os ésteres IP6 e IP5.
O conteúdo de substrato exerce um efeito importante sobre a atividade da fitase, e sua ação está na dependência da presença do substrato e sua concentração. Apesar do aumento da concentração de substrato não causar um aumento indefinido na atividade enzimática, a pouca presença do substrato pode diminuir sua atividade, já que o contato físico entre enzima e substrato é necessário. O Km da enzima (Figura 6) descreve a habilidade que uma enzima tem para manter sua atividade quando a quantidade de substrato em solução é limitante. De maneira geral, as fitases bacterianas apresentam Km mais baixo que as fitases fúngicas (nesse caso quanto menor melhor), e estão surgindo algumas fitases E. coli geneticamente modificadas onde se está incrementando ainda mais a afinidade ao substrato, e adicionalmente se incrementando a resistência às proteases endógenas e pH baixo.
Outro ponto de grande relevância é a termo-estabilidade das enzimas. De acordo com Ushasree (2011) a fitase ideal é aquela que é intrinsecamente termotolerante, e a engenharia genética tem ajudado cada vez mais a alcançar a fitase ideal (do ponto de vista de termo-tolerância e eficiência catalítica).
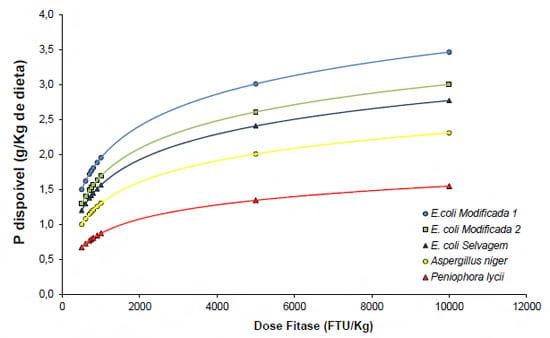
Tomando-se em conta todos os fatores acima expostos é de se supor que diferentes fitases tenham liberações de fósforo distintas. Selle e Ravindran (2007), em um artigo de revisão bibliográfica sobre fitases, demonstraram que existe muita variação na liberação de fósforo pelas fitases, e isto pode estar correlacionado com a metodologia de avaliação empregada no experimento. Dados internos da AB Vista têm demonstrado que as E.coli fitases (especialmente as duas gerações de E.coli fitases geneticamente modificadas) têm uma maior capacidade de liberação de fósforo do fitato que as fitases fúngicas (Figura 7).
Figura 6 – Efeito da concentração de substrato na velocidade de reação catalisada enzimaticamente
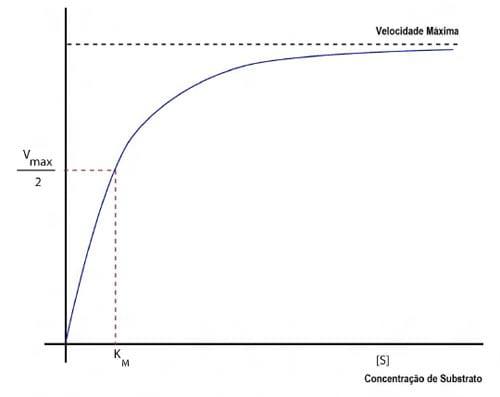
Figura 7 – Efeito da dose-resposta sobre a disponibilização de fósforo em dietas recebendo a inclusão de diferentes fitases
“Superdosagem” de fitase e seus efeitos extra-fosfóricos
Por todo o efeito danoso que o fitato exerce sobre a digestibilidade de nutrientes em aves, estão sendo feitos experimentos ao redor do mundo com doses maiores de fitases, além dos 500FTU’s convencionais.
Apesar de ser um tema que ainda está em desenvolvimento, principalmente quando olhamos nas possibilidades de aplicação comercial, os estudos iniciais com doses mais elevadas de fitase datam do inicio da década de 70. Nelson, um dos pioneiros nos estudo com fitases, conduziu um estudo empregando doses elevadas de fitase (0, 950, 1900, 3800, 7600FTU/Kg de uma fitase de Aspegillus ficuum) no desempenho de frangos de corte. Os resultados mostraram que a maior dosagem (7600FTU/Kg) teve a melhor resposta após 21 dias de idade das aves, onde o ganho de peso e cinzas ósseas incrementaram 31 e 59%, respectivamente, quando comparadas às aves que não receberam fitase.
Em estudos posteriores (Auspurger e Baker, 2004; Brana et al., 2006; Cowieson et al., 2004) se comprovou o achado de Nelson, onde nesses estudos se utilizou até 24000FTU/Kg, e se verificou incrementos significativos no desempenho dos animais.
Selle e Ravindran (2007) mencionam que estratégias devem ser adotadas para diminuir o conteúdo de fitato ingerido pelos animais, seja pela desfitinização das matérias-primas (principalmente quando falamos de farelos de oleaginosas), pela diminuição do conteúdo de fitato nos grãos vegetais através da modificação genética, ou pela utilização de fitases mais eficientes na nutrição animal.
Recentemente Cowieson et al. (2011) publicou um artigo abordando a estratégia de “Superdosagem” de fitase, especialmente focando as fitases provenientes de E.coli e Aspergillus niger, e concluíram que existem oportunidades muito interessantes na melhoria da conversão alimentar e ganho de peso em aves e suínos, inclusive podendo melhorar o rendimento de carcaça dos animais. Bedford (2012) enumerou recentemente alguns dos benefícios da utilização da “Superdosagem” de fitase em aves (frangos, poedeiras, perus e patos).
Bedford et al. (2011) demostraram efeitos bastante benéficos sobre o ganho de peso de perus quando altas dosagens de fitase foram empregados. Os resultados desse estudo demonstram que apenas 500FTU’s de uma E.coli fitase geneticamente modificada foram capazes em compensar uma diminuição em 0,15% no fósforo disponível e 0,15% no cálcio total (dieta controle negativo adicionados de 500FTU/Kg), apresentando resultados similares aos animais alimentados com a dieta controle positivo. Adicionalmente, animais recebendo 2000FTU/Kg apresentaram um maior ganho de peso (3,5% superior) quando comparados aos animais recebendo a dieta controle positivo.
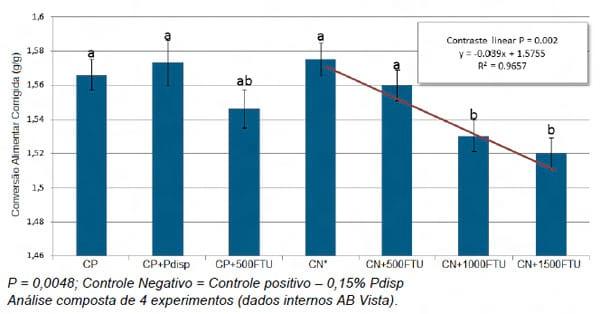
Dados internos da AB Vista demonstram um benefício bastante interessante da utilização da “Superdosagem” de uma E.coli geneticamente modificada, de segunda geração, sobre a conversão alimentar de frangos de corte (Figura 8).
Dois outros fatores podem ser de grande importância quando da utilização do conceito de “Superdosagem”. O primeiro deles é o potencial aproveitamento da assimilação de mio-inositol. Karadas et al. (2010), em estudo onde utilizaram até 12.500FTU/Kg de uma E. coli fitase geneticamente modificada, verificaram um aumento nos níveis de vitamina A, vitamina E e Coenzima Q10. Isso porque o inositol desempenha um importante papel no transporte de lipídeos, e a Coenzima Q10, um importante antioxidante, pode auxiliar nas funções mitocondriais e assim trazer benefícios não apenas para o metabolismo energético, mas também na transferência de nutrientes para o embrião quando da formação da gema. Walk et al. (2012) mencionam ainda a possibilidade de empregar “Superdosagem” de uma E. coli geneticamente modificada para, além de melhorar o desempenho dos animais, poder reduzir a inclusão de calcário nas dietas, já que esse ingrediente causa diversas reações fisiológicas no organismo prejudicando a digestibilidade de nutrientes. No estudo realizado por esses autores a utilização de 5000FTU/Kg em dietas contendo apenas 0,64% de cálcio total foi capaz em melhorar a conversão alimentar em 50g (1,356 vs 1,306) e o ganho de peso em 10g (462 vs 472) quando comparadas a aves recebendo a dieta controle positivo (1,03% de cálcio total sem a adição de fitase). Adicionalmente, os autores verificaram um incremento estatisticamente significativo na digestibilidade ileal de aminoácidos (Arg, Val, Iso, Leu, Phe, Lys, Asp, Ser, Glu, Pro, Gly, Ala, Tyr) e uma tendência de incremento na digestibilidade da proteína bruta, quando da utilização de “Superdosagem” de fitase, sendo que a maior quantidade de cálcio (via calcário) parece afetar a digestibilidade da proteína bruta.
Figura 8 – Efeito de diferentes dosagens de uma E.coli geneticamente modificada sobre a conversão alimentar corrigida de frangos de corte aos 42 dias de idade
Conclusões
Nos últimos anos passamos a compreender melhor os efeitos anti-nutricionais do fitato e seus reflexos na utilização de nutrientes pelos animais. Este melhor conhecimento levou a uma série de novas pesquisas no intuito de elucidar os benefícios da redução do fitato, pelo uso da “superdosagem” de fitase, sobre os parâmetros de desempenho de aves. Apesar do mecanismo pelo qual as “superdosagens” de fitase incrementam os parâmetros de desempeno não estar completamente elucidado, está claro que existe um incremento da digestibilidade e do status anti-oxidante dos animais. Os dados são realmente promissores, e sugerem que o futuro das fitases não mais será a liberação de fósforo apenas, mas a busca incessante pela enzima mais hábil em rapidamente hidrolisar o fitato buscando assim os efeitos extra-fosfóricos advindos da utilização da chamada “superdosagem” de fitase.
Referências Bibliográficas
Auspurger, N.R.; Baker, D.H. (2004) High dietary phytase levels maximize phytate phosphorus utilization but do not affect protein utilization in chicks fed phosphorus or amino acid deficient diets. Journal of Animal Science, 82: 1100-1107.
Bedford, M.R.; Wyatt, C.; Knox, A.; Santos, T.T. (2011), Turkey performance between 0 and 84 days when diets supplemented with high doses of phytase, International Poultry Symposium, Atlanta.
Bedford, M.R. (2012) Alternate uses of phytases – Superdosing. Asian Poultry Magazine, p. 8-11.
Brana, D.V.; Ellis, M.; Castandeda, E.O.; Sands, J.S. (2006), Effect of novel phytase on growth performance, bone ash, mineral digestibility in nursery and grower-finisher pigs. Journal of Animal Science, 84: 1839-1849.
Coelho, C.M.M.; Santos, J.C.P.; Tsai, S.M.; Vitorello, V.A. (2002) Seed phytate content and phosphorus uptake and distribution in dry bean genotypes. Brazilian Journal of Plant Physiology, 14: 51-58.
Cowieson, A.J.; Acamovic, T.; Bedford, M.R, (2004), The effect of phytase and phytic acid on the endogenous loss from broiler chickens. British Poultry Science, 45: 101 - 108.
Cowieson, A.J.; Cowieson, N.P. (2011), Phytate and the thermodynamics of water. Australian Poultry Symposium, Page 11.
Cowieson, A.J.; Ravindran, V (2007), Effect of phytic acid and microbial phytase on the flow and amino acid composition of endogenous protein at the terminal ileum of the growing broiler chicken. British Journal of Nutrition, 98: 45-752
Cowieson, A.J.; Wilcock, P.; Bedford, M.R. (2011) Super-dosing effects of phytase in poultry and other monogastrics. World’s Poultry Science Journal, 67: 225-236.
Harland, B.F.; Morris, E.R. (1995), Phytate – A good or bad food component? Nutrition Research, 15: 733-754
Igbasan, F.A.; Männer, K; Miksch, G.; Borriss; R.; Farouk, A.; Simone, O. (2000) Comparative studies on the in vitro properties of phytases from various microbial origins, Archiv für Tierernaehrung, 53:353-373.
Karadas, F.; Pirgozliev, V.; Pappas, A.C.; Acamovic, T.; Bedford, M.R. (2010), Effects of different dietary phytase activities on the concentration of antioxidants in the liver of growing broilers Journal of Animal Physiology and Animal Nutrition. 94: 519-526.
Kies,A.K.; De Jonge, L.H; Kemme,P.A.; Jongbloed, A.W. (2006) Interaction between Protein, Phytate, and Microbial Phytase. In Vitro Studies. Journal of Agricultural and Food Chemistry, 54:1753–1758.
Krabbe, E.L. (2012) Perspectivas quanto ao desenvolvimento de enzimas para uso na nutrição de aves. XI Seminário Internacional de Aves e Suínos, São Paulo, Brazil, CD ROM.
Luttrell, B.M. (1993) The biological relevance of the binding of calcium ions by inositol phosphates. The Journal of Biological Chemistry, 268: 1521-1524.
Nelson, T.S. (1984) Available calcium for poultry. Proceedings of the Florida Nutrition Conference for Feed Manufacturers, Orlando, Florida, pp. 1-6.
Onyango, E.M.; Bedford, M.R.; Adeola, O. (2005) Phytase activity along the digestive tract ofthe broiler chick: A comparative study ofan Escherichia coli-derived and Peniophora lycii phytase. Canadian Journal of Animal Science, 85: 61-68.
Prata, R.; Batie, C.; Betts, S.; Basu, S.S. (2007) XX Congreso Latinoamericano de Avicultura, Porto Alegre, Brazil, p.13-15.
Rostagno, H.S.; Albino, L.F.T.; Donzele, J.L.; Gomes, P.C.; Oliveira, R.F.M.; Lopes, D.C.; Ferreira, A.S.; Barreto, S.L.T.; Euclydes, R.F. Tabelas brasileiras para aves e suínos – Composição de alimentos e requerimentos nutricionais. 3. ed. Visconde do Rio Branco, MG: Suprema, 2011. v. 01. 259 p.
Santos, T.T. (2012) Phytate: anti-nutrient for poultry and swine. Feedstuffs, 84:1-3.
Selle, P.H.; Ravindran, V. (2007) Microbial phytase in poultry nutrition. Animal Feed Science and Technology, 135:1-41.
Tamim, N.M.; Angel, R. (2003) Phytate phosphorus hydrolysis as influenced by dietary calcium and micro-mineral source in broiler diets. Journal of Agriculture and Food Chemistry, 51: 4687-4693.
Tamim, N.M.; Angel, R.; Christman, M. (2004) Influence of dietary calcium and phytase on phytate phosphorus hydrolysis in broiler chickens. Poultry Science, 83:1358–1367.
Ushasree, M.V.; Sumayya, H.B.V.S.; Pandey, A. (2011) Adopting structural elements from intrinsically stable phytase – a promising strategy towards thermostable phytases. Indian Journal of Biotechnology, 10:458-467.
Wadt, G.R.; Santos, T.T.; Gomes, G.A.; Cowieson, A.J.; Bedford, M.R. Total and phytic phosphorus on corn samples from different regions in Brazil. Proceedings of Congresso Latino Americano de Nutrição Animal. Estância de São Pedro, 4:7-8.
Walk, C.L., Bedford, M.R., McElroy, A.P. (2012) Influence of limestone and phytase on broiler performance, gastrointestinal pH, and apparent ileal nutrient digestibility. Poultry Science, 91:1371–1378.
Tópicos relacionados
Autores:

Junte-se para comentar.
Uma vez que se junte ao Engormix, você poderá participar de todos os conteúdos e fóruns.
* Dados obrigatórios
Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.
Criar uma publicação
18 de marzo de 2015
Olá Gilson, excelente artigo! Vc teria essas referências em pdf?

Opta Alimentos e Insumos
2 de marzo de 2015
Gilson, grato pelo belo artigo!
Como é sua percepção na interação de duas fitases, uma 6-fitase e outra 3-fitase em dosagem regular e em super dosagem, podemos ter sinergias?
Cordialmente,
Luis Luna





Você pode estar interessado em
.jpg&w=3840&q=75)