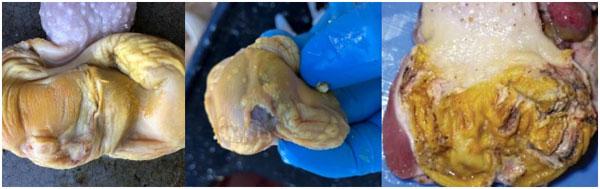
Descrição da síndrome de erosão da moela em aves comerciais no Brasil
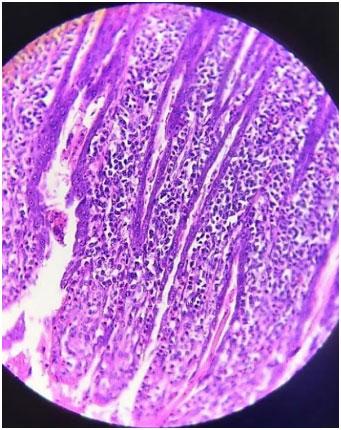
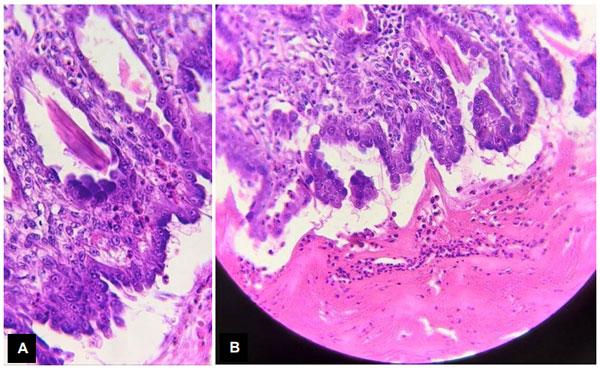
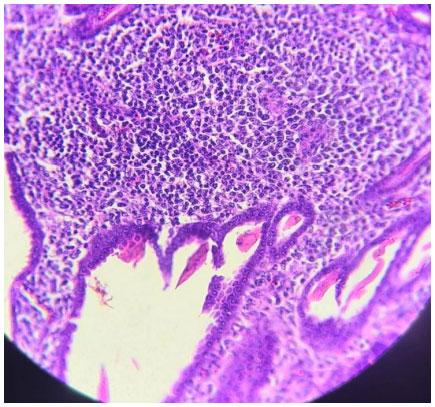
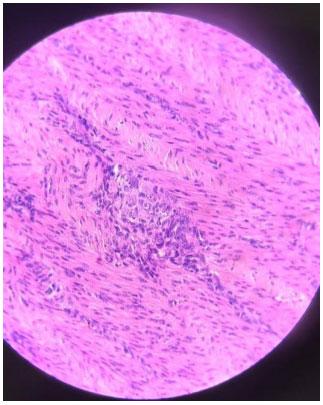
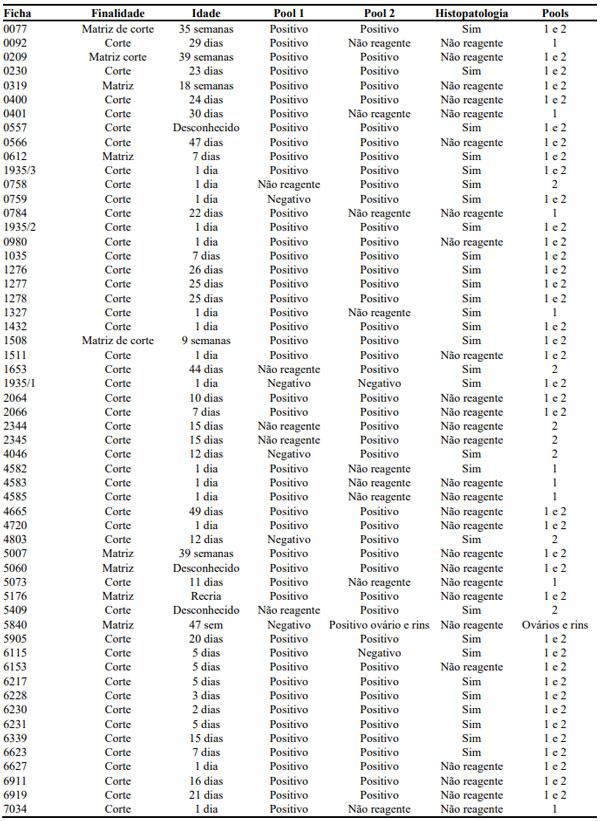
Infecções adenovirais são comuns em aves comerciais, principalmente em sistemas de criação intensificada e já foram reportadas em diferentes locais, como: Ásia, África, América do Norte, América do Sul e Europa. As doenças envolvendo os Aviadenovírus (FAdVs), incluem a hepatite por corpo de inclusão (HCI), síndrome da hepatite-hidro pericárdio, (SHH) e outras condições afetando o sistema respiratório e digestivo das aves, além da Síndrome de Erosão de Moela (SEM). Esta publicação objetiva a caracterização da SEM como condição emergente no Brasil. Foram analisadas, entre setembro de 2021 e maio de 2022, 168 aves em 56 casos, sendo estas tanto de corte como de reprodução. Foram conduzidos exames de PCR para identificar o DNA viral e 28 exames histopatológicos com alguns destes revelando presença de corpos de inclusão virais no ventrículo, além de quadros inflamatórios típicos envolvendo infiltrado linfoplasmohistiocítico. Foram encontrados outros processos inflamatórios em outros órgãos, como o pró-ventrículo. O PCR evidenciou positividade para o Aviadenovírus. Com base nos achados do presente estudo, caracterizou-se a presença da SEM no Brasil. Enfatizamos a importância do sequenciamento do vírus, especialmente do tipo 1, como próximo passo a ser seguido.
Palavras-chave: Adenovirus, aviadenovirus, corpos de inclusão, FAdV, histopatologia
Publicado originamente na revista eletrônica PUBVET, em agosto de 2022. Acesso disponível em:http://www.pubvet.com.br/artigo/10070/descriccedilatildeo-da-siacutendrome-de-erosatildeo-da-moela-em-aves-comerciais-no-brasil
Absalón, A. E., Morales-Garzón, A., Vera-Hernández, P. F., Cortés-Espinosa, D. V, Uribe-Ochoa, S. M., García, L. J., & Lucio-Decanini, E. (2017). Complete genome sequence of a non-pathogenic strain of Fowl Adenovirus serotype 11: Minimal genomic differences between pathogenic and nonpathogenic viruses. Virology, 501, 63–69. https://doi.org/10.1016/j.virol.2016.11.006.
De la Torre, D., Nunez, L. F. N., Parra, S. H. S., Astolfi-Ferreira, C. S., & Ferreira, A. J. P. (2018). Molecular characterization of fowl adenovirus group I in commercial broiler chickens in Brazil. VirusDisease, 29(1), 83–88. https://doi.org/10.17163/lgr.n28.2018.06.
Dezengrini, R., Weiblen, R., & Flores, E. F. (2007). Soroprevalência das infecções por parvovírus, adenovírus, coronavírus canino e pelo vírus da cinomose em cães de Santa Maria, Rio Grande do Sul, Brasil. Ciência Rural, 37(1), 183–189.
Duncan, J. R., & Prasse, K. W. (1982). Patologia clínica veterinária. UFSM.
Fitzgerald, S. D., Rautenschlein, S., Mahsoub, H. M., Pierson, F. W., Reed, W. M., & Jack, S. W. (2019). Adenovirus infections. Diseases of Poultry, 321–363. https://doi.org/10.1002/9781119371199.ch9.
Grafl, B., Garcia-Rueda, C., Cargill, P., Wood, A., Schock, A., Liebhart, D., Schachner, A., & Hess, M. (2018). Fowl aviadenovirus serotype 1 confirmed as the aetiological agent of gizzard erosions in replacement pullets and layer flocks in Great Britain by laboratory and in vivo studies. Avian Pathology, 47(1), 63–72. https://doi.org/10.1080/03079457.2017.1367364.
Grafl, B., Liebhart, D., Günes, A., Wernsdorf, P., Aigner, F., Bachmeier, J., & Hess, M. (2013). Quantity of virulent fowl adenovirus serotype 1 correlates with clinical signs, macroscopical and pathohistological lesions in gizzards following experimental induction of gizzard erosion in broilers. Veterinary Research, 44(1), 1–8.
Hess, M. (2013). Aviadenovirus infections. Diseases of Poultry, 290–300.
Luna, L. G. (1971). Manual of histologic staining methods of the Armed Forces Institute of Pathology. Pathology, 3(3), 249. https://doi.org/10.1016/s0031-3025(16)39410-7.
Matczuk, A. K., Niczyporuk, J. S., Kuczkowski, M., Woźniakowski, G., Nowak, M., & Wieliczko, A. (2017). Whole genome sequencing of Fowl aviadenovirus A-a causative agent of gizzard erosion and ulceration, in adult laying hens. Infection, Genetics and Evolution, 48, 47–53. https://doi.org/10.1016/j.meegid.2016.12.008.
Mettifogo, E., Nuñez, L. F. N., Parra, S. H. S., Astolfi-Ferreira, C. S., & Ferreira, A. J. P. (2014). Fowl adenovirus Group I as a causal agent of inclusion body hepatitis/hydropericardium syndrome (IBH/HPS) outbreak in brazilian broiler flocks. Pesquisa Veterinária Brasileira, 34, 733–737.
Meulemans, G., Boschmans, M., Van den Berg, T. P., & Decaesstecker, M. (2001). Polymerase chain reaction combined with restriction enzyme analysis for detection and differentiation of fowl adenoviruses. Avian Pathology, 30(6), 655–660. https://doi.org/10.1080/03079450120092143.
Mirzazadeh, A., Grafl, B., Abbasnia, M., Emadi-Jamali, S., Abdi-Hachesoo, B., Schachner, A., & Hess, M. (2021). Reduced performance due to adenoviral gizzard erosion in 16-day-old commercial broiler chickens in Iran, confirmed experimentally. Frontiers in Veterinary Science, 8, 635186. https://doi.org/10.3389/fvets.2021.635186.
Naranjo, L. F. N. (2012). Isolamento e propagação de Astrovírus, Adenovírus, Coronavírus, Parvovírus, Rotavírus e Reovírus de aves comerciais com problemas entéricos em ovos embrionados. Universidade de São Paulo.
Ono, M., Okuda, Y., Yazawa, S., Imai, Y., Shibata, I., Sato, S., & Okada, K. (2003). Adenoviral gizzard erosion in commercial broiler chickens. Veterinary Pathology, 40(3), 294–303.
Peterson, T. S., Spitsbergen, J. M., Feist, S. W., & Kent, M. L. (2011). Luna stain, an improved selective stain for detection of microsporidian spores in histologic sections. Diseases of Aquatic Organisms, 95(2), 175–180. https://doi.org/10.3354/dao02346.
Schachner, A., Grafl, B., & Hess, M. (2021). Spotlight on avian pathology: fowl adenovirus (FAdV) in chickens and beyond–an unresolved host-pathogen interplay. Avian Pathology, 50(1), 2–5. https://doi.org/10.1080/03079457.2020.1810629.
Schachner, A., Matos, M., Grafl, B., & Hess, M. (2018). Fowl adenovirus-induced diseases and strategies for their control–a review on the current global situation. Avian Pathology, 47(2), 111– 126. https://doi.org/10.1080/03079457.2017.1385724.
Slaine, P. D., Ackford, J. G., Kropinski, A. M., Kozak, R. A., Krell, P. J., & Nagy, É. (2016). Molecular characterization of pathogenic and nonpathogenic fowl aviadenovirus serotype 11 isolates. Canadian Journal of Microbiology, 62(12), 993–1002. https://doi.org/10.1139/cjm-2016-0297.
Tanimura, N., Nakamura, K., Imai, K., Maeda, M., Gobo, T., Nitta, S., Ishihara, T., & Amano, H. (1993). Necrotizing pancreatitis and gizzard erosion associated with adenovirus infection in chickens. Avian Diseases, 37(2), 606–611. https://doi.org/10.2307/1591697.
Thomson, R. G. (1990). Patologia veterinária especial (Vol. 1). Manole.
Vera-Hernández, P. F., Morales-Garzón, A., Cortés-Espinosa, D. V, Galiote-Flores, A., García-Barrera, L. J., Rodríguez-Galindo, E. T., Toscano-Contreras, A., Lucio-Decanini, E., & Absalón, A. E. (2016). Clinicopathological characterization and genomic sequence differences observed in a highly virulent fowl Aviadenovirus serotype 4. Avian Pathology, 45(1), 73–81. https://doi.org/10.1080/03079457.2015.1125443.






