Sêneca, um dos mais célebres filósofos do império romano, declarou: “Sorte é o que acontece quando o preparo se encontra com a oportunidade”. No decorrer do nosso trabalho percebemos a veracidade deste pensamento.
Adentrar a área de modelagem foi uma decisão difícil, pois a matemática assombra a muitos desde a meninice. Mas, quando ousamos, descobrimos que esse fantasma pode ser nosso camarada (Sampaio, 2002). Foi exatamente o que aconteceu, quando empenhávamos no aperfeiçoamento da planilha PPM.
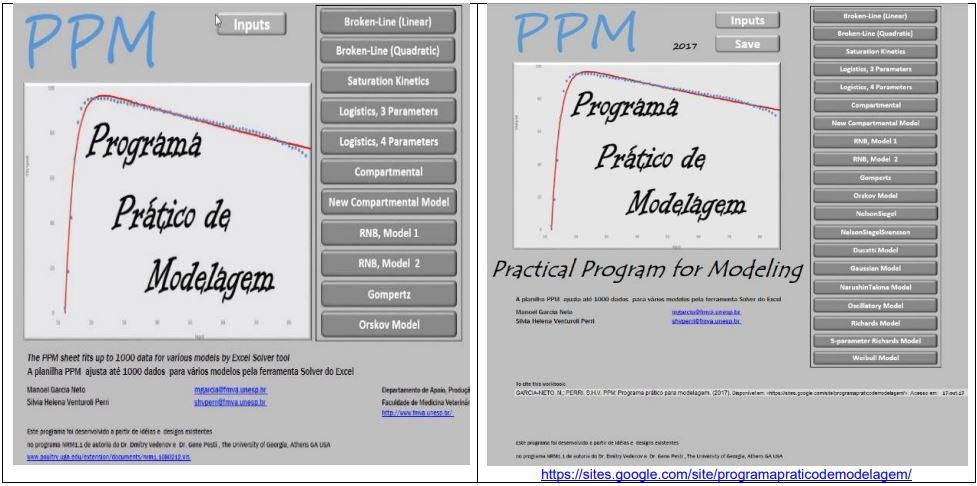
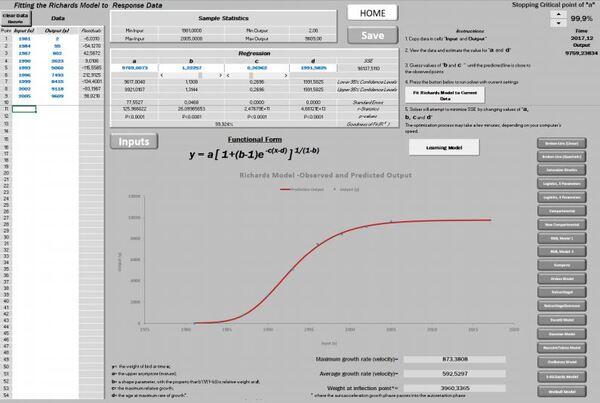
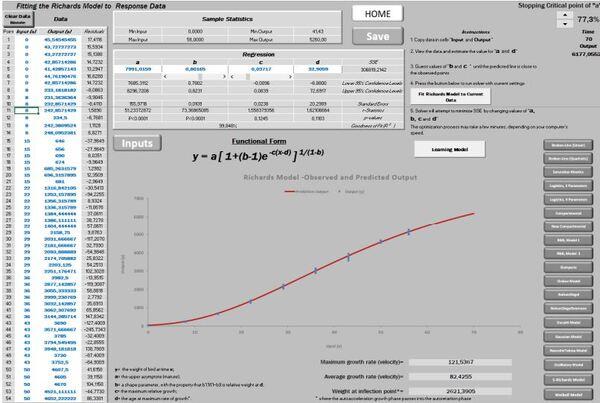
No início apenas desejávamos encontrar um programa livre que ajustasse a tão conhecida curva de Gompertz. Deparamos com um programa em Excel de autoria de Petr Klemera e Stanislav Doubal, que gentilmente nos enviou uma cópia. Continuamos à procura de algo que suprisse a nossa necessidade. Então, acessamos o programa NRM1.1 de Verdenov e Pesti que é disponibilizado livremente, entretanto há o limite de apenas 100 entradas. Como nossos dados ultrapassavam esse limite, percebemos que as duas planilhas já citadas, não viabilizavam rodar nossos números. Assim, conhecendo a capacidade do suplemento solver do Excel tivemos a oportunidade de iniciar uma nova planilha com 1000 dados de entrada (Kemmer & Keller, 2010). Nasce, assim, a planilha PPM, ajustando inicialmente 11 modelos, e atualmente 20 (Figuras 1 e 2).
Figura 1.
Planilhas livres para ajustes de curvas
Figura 2. Planilha PPM para ajustes de curvas
Certamente, nada cresce na natureza sem atingir um limite (Karkach, 2006). Logo, surge a questão: Qual será o limite para a planilha PPM? Relembrando que o objetivo inicial era um programa que ajustasse a curva de Gompertz.
Ao participarmos de uma banca de Doutoramento em Pirassununga com o Doutor Douglas E. de Faria (http://www.teses.usp.br/teses/disponiveis/74/74131/tde-03112016- 140320/publico/DO8193265COR.pdf), foi nos apresentado a curva de Richards e seus atributos, o que muito nos fascinou; mas ocorrendo um habitual inconveniente “não convergia”, possivelmente devido a problemas de compatibilidades numéricas (Yin et al., 2003; Karkach, 2006).
Diante desse novo desafio, começamos a buscar caminhos para a curva de Richards convergir. Há muito tempo, outros pesquisadores, já buscavam solucionar esse enigma; como Causton (1969) ao propor “A computer program for fitting the Richards function”. Isso posto, cabe um alerta aos futuros pesquisadores, não se debrucem apenas à Literatura “dos últimos dez anos”, pois muitas vezes a resposta não estará na bibliografia mais atual (Fisher et al., 1996ab).
Jorgensen (1980) de forma cuidadosa e genial propõe o ajuste da curva de Richards. Analogicamente, podemos entender a proposta dele, trazendo a figura do malabares que deseja lançar sempre todos os pinos ao ar, mas sem sucesso. Entretanto, se ele modestamente começar a jogar dois pinos, depois três e finalmente os quatro, certamente terá maior possibilidade de obter sucesso.
Assim, iniciamos ao ajuste da curva de Richards que segundo Jorgensen deveríamos começar de forma comedida os parâmetros a, b, c, e d, mas não todos ao mesmo tempo. Provavelmente, essa é a razão do grande insucesso no ajuste da curva de Richards, encontrados na literatura (Freitas et al., 1984; Zach, 1988; Braccini-Neto et al., 1996; Birch, 1999; Oliveira et al., 2000; Venugopalan & Shamasundaran, 2003; Vitezica et al., 2010; Porter et al., 2010; Drumond et al., 2013; Faridi et al., 2014, Veloso et al., 2015, 2016) mesmo utilizando programas sofisticados como o SAS (retomando a analogia: jogam todos os quatro pinos/parâmetros de uma só vez). Por isso, voltamos a afirmar: as respostas que desejamos já podem ter sido esclarecidas há muito tempo, para encontrá-las há necessidade de realizar uma boa revisão que poderá ultrapassar os “últimos dez anos de literatura”.
Aplicando esse princípio da não avidez em querer ajustar os quatro parâmetros de uma vez só, obtivemos o tão almejado sucesso de ajustar a curva de Richards. Entretanto, ainda há particulares ajustes e situações que requerem mais paciência. Para esses casos mais melindrosos foi disponibilizado um botão específico “Learning Model “.
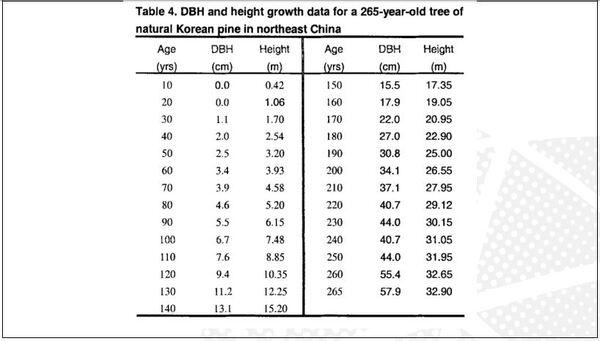
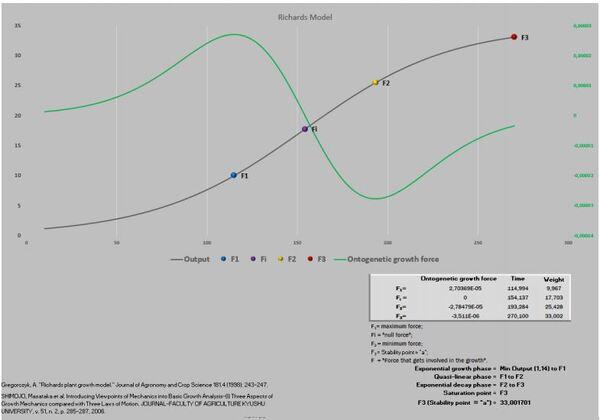
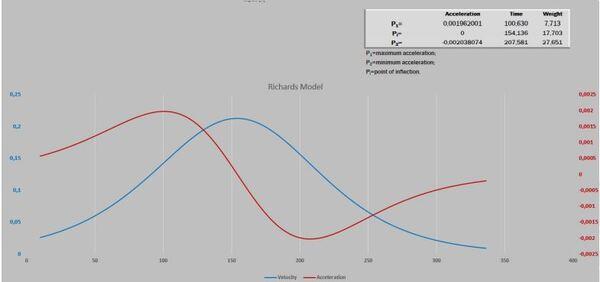
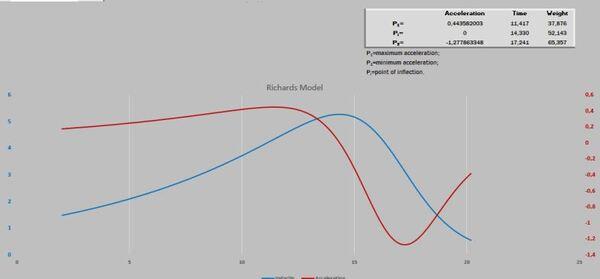
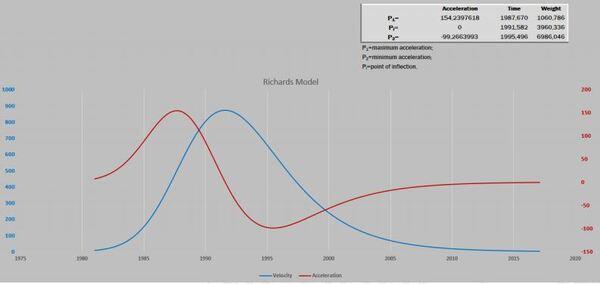
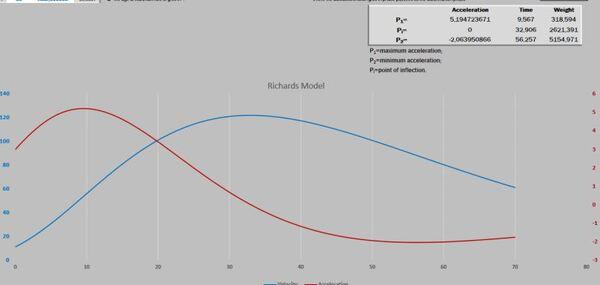
Visando melhorar a interpretação das curvas, acrescentamos o gráfico de resíduos. Após isso, deparamos com um novo desafio proposto por Gregorczyk (1991), desdobrar a curva de crescimento em sua primeira derivada (velocidade) e segunda derivada (aceleração), aspirando encontrar os pontos definidos na curva P1, Pi e P2. A partir desses pontos teriam três fases na curva de crescimento: a fase exponencial, a fase linear (P1 até P2) e a fase de maturação. É interessante observar que os pontos P1 e P2 são simétricos ao Pi. A razão disso é que a curva analisada era a logística que tem seu ponto de inflexão exatamente em 50% do valor de sua assíntota (Figura 3).
Figura 3. Pontos críticos definidos por derivada.
Trabalhos mais recentes apresentam uma visão mecanicista aos modelos de crescimentos e abordam uma visão ontogênica ao destacar fases críticas durante a trajetória do crescimento (Forbes & Lopez, 1989; Wu et al., 2004; Hall et al., 2013), derivando a curva de crescimento para obter os mesmos três pontos críticos P1, Pi e P2
A fim de mostrar a importância desses pontos, Wu et al. (2004) apresentaram um exemplo esclarecedor ao descrever as três fases do câncer: o crescimento inicial exponencial, a fase intermediária linear, e por fim, o declínio do crescimento do tumor. Ao se conhecer tal trajetória seria possível definir com mais precisão o início dos sintomas clínicos e o momento da manifestação dos mais sérios problemas.
Desta forma, convictos de que a curva de crescimento permite revelar mais compreensão por suas derivadas, e principalmente que o modelo de Richards define a curvaefetivamente mais adequada para os dados analisados, tudo isso fez com que iniciássemos novos ajustes ao PPM (Marinakis, 2012).
Nesse momento estávamos diante de um extremo desafio matemático: derivar a curva de Richards em sua primeira e segunda derivada. Felizmente, nos deparamos com recursos livres na internet que de uma forma magistral acomodaram nosso “fantasma”, não somente para a curva de Richards, como também, para Gompertz, Logísticas (2 e 3) e Weibull.
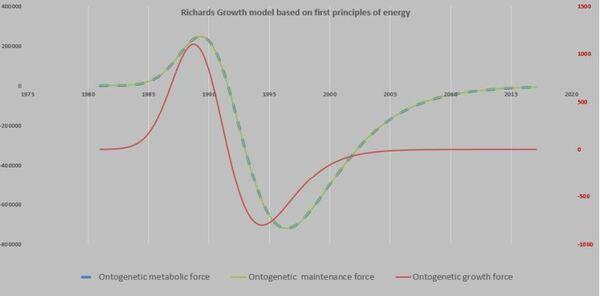
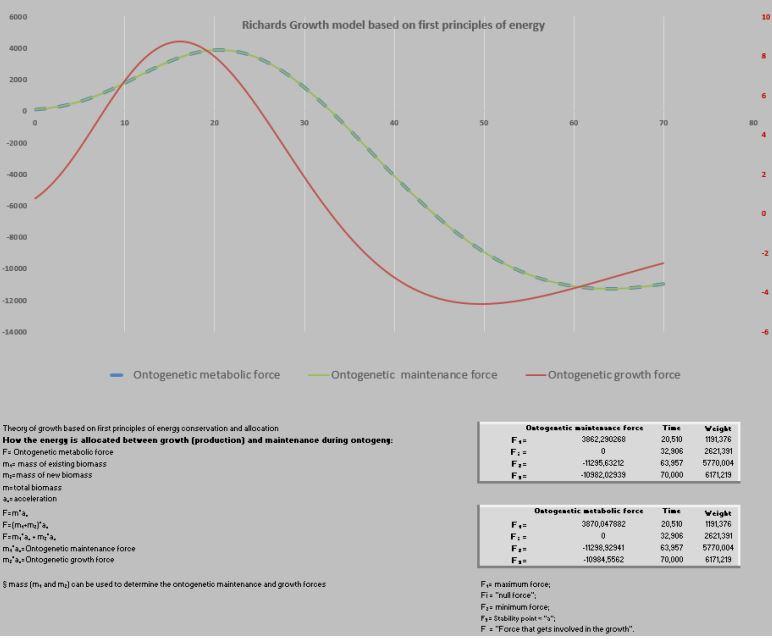
Então, ficamos animados por encontrar o caminho das derivadas e suas aplicações práticas para avaliar a influência favorável ou não da nutrição, genética e ambiência em relação às curvas de crescimento. Nesse momento do PPM surge uma nova proposta dizendo que seria possível aplicar uma visão mecanicista, com base nas leis de Newton (Shimojo et al. 2006).
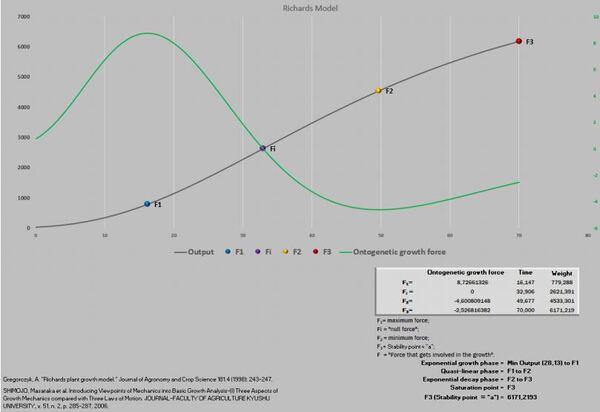
Shimojo et al. (2006) passam a definir novos termos como “growth force” e que F= m.a “looks like force that gets involved in the growth”.
Daí a necessidade de retomarmos o pensamento de Sêneca: “Sorte é o que acontece quando a capacidade se encontra com a oportunidade”, podemos considerar que tivemos “sorte”, pois tínhamos a capacidade da planilha PPM em definir a aceleração (segunda derivada da curva de crescimento) e conhecer a “massa” em qualquer ponto da curva de crescimento; e a oportunidade de aplicar a proposta de Shimojo et al. (2006) na planilha PPM.
Entramos, então, numa nova área que visa contemplar as “forças” e suas trajetórias. Alguns autores já se debruçaram sobre esse tema (Von Bertalanffy, 1957; Forbs & Lopes, 1989; West et al. 2001; Makarieva et al., 2004; West et al., 2004; Moses et al., 2008; Dumas et al., 2010; Hook et al., 2011; Hall et al.,2013; Griebeler et al., 2013; Shi et al., 2014; Mahanta & Borah, 2014; Maino & Kearney, 2015; Martyshev & Terentivev, 2015).
Após essa revisão, definimos ser conveniente batizar a “força do crescimento/ growth force” como “força ontogênica metabólica” e seu desdobramento em “força ontogênica do crescimento” e “força ontogênica de manutenção”, a saber:
F = m.a;
correspondendo F= força ontogênica metabólica;
m = massa mensurada em cada tempo da curva;
a = aceleração da massa correspondente ao aferir a massa.
Como o princípio da conservação da energia deve ser observado e não pode ser violado (Makarieva et al.,2004; Shi et al., 2014), inferimos que a massa poderia ser dividida em m1 e m2: m1= massa da já existente biomassa e m2= massa da nova biomassa
Assim sendo:
F=m.a
F= (m1 + m2).a
F= m1.a + m2.a
Significando:
m1.a = força ontogênica de manutenção da massa já existente, entretanto em
m2.a = força ontogênica do crescimento, ou seja, a energia destinada à síntese de nova biomassa que efetivamente faz o animal crescer dia a dia até atingir seu máximo desenvolvimento (assíntota) (Shi et el., 2014).
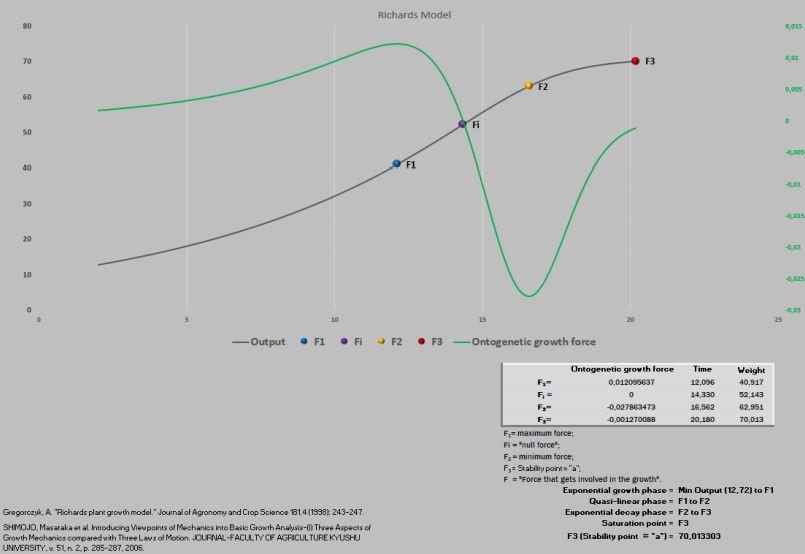
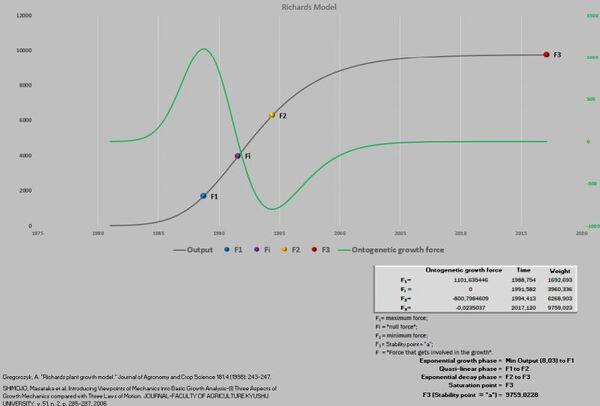
Essas referidas novidades exigiram uma nova abordagem da planilha PPM, visando acomodar esses novos pontos que foram definidos como F1, Fi, F2 e F3.
Massa ao nascimento até F1 = fase exponencial do crescimento.
F1 até F2 = fase quase-linear do crescimento.
F2 até F3 = fase exponencial decrescente.
F3 = ponto de saturação (final do crescimento).
Como aplicação prática para elucidar essas novidades serão ajustadas curvas a fim de avaliar a ontogenia do crescimento em árvores (altura), em humanos (kg), de aids (pessoas) e, finalmente, em frangos (gramas).
Exemplo 1 – Predição de altura de pinheiros
Propositadamente escolhemos o trabalho de Zhao-gang & Feng-ri (2003), em virtude desses autores aferirem o crescimento em “metros”, o que segundo Shi et al. (2014) seria compatível de ser ajustado por curvas sigmoides. Surpreendentemente, os dados dessa pesquisa abrangem um período compreendido de 265 anos. Por questões óbvias, a massa em kg não seria a melhor opção (Karkach, 2006), mas sim a altura das árvores (Figura 4).
Figura 4. Altura de pinos conforme a idade (anos).
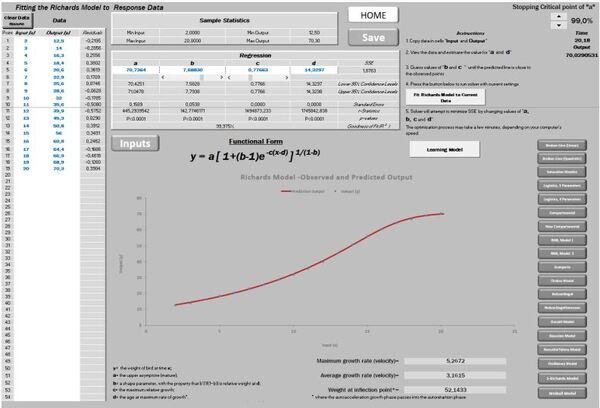
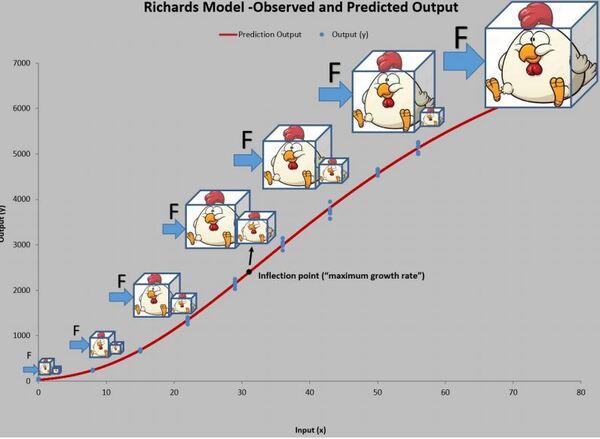
Dessa forma, ao rodar os dados na planilha PPM, obtém-se um ajuste de R2= 99,93 com crescimento máximo de 21,24 cm/ano. E ainda, uma altura no ponto de inflexão (ápice da velocidade de crescimento) de 17,70 metros.
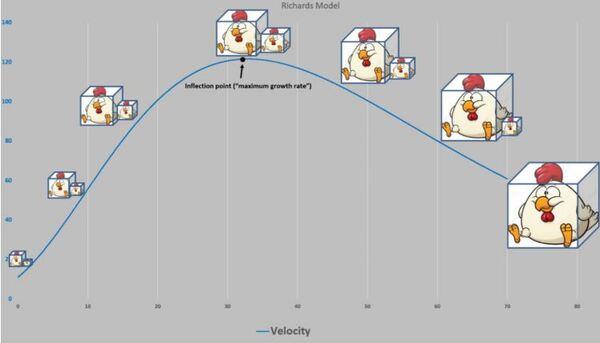
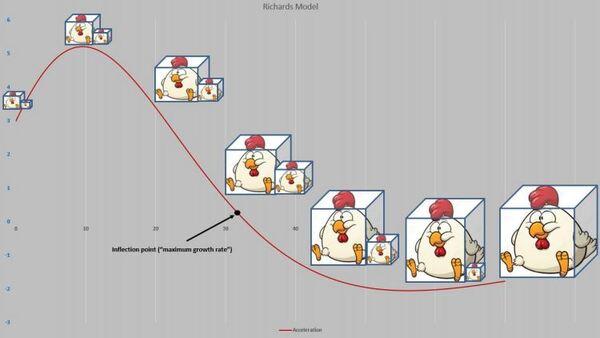
A Figura 5 apresenta as curvas de velocidade e aceleração e os pontos tradicionais P1, Pi e P2 (Gregorczyk, 1998). Curiosamente é a equidistância da altura 7,7-17,7 (+10) e 17,7 – 27,7(+10), lembrando uma curva logística (Ponto de inflexão em 50% da assíntota).
Figura 5. Curva ajustada pela planilha PPM e subsequente primeira e segunda derivadas (velocidade e aceleração).
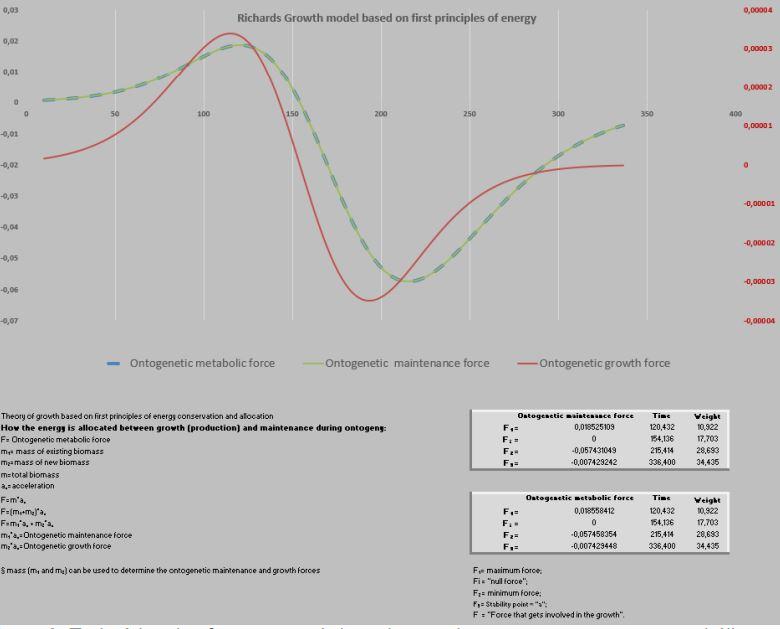
Agora surge a parte mais desafiadora, avaliar as forças metabólicas envolvidas no crescimento de pinos por 265 anos (Figura 6), e a “massa” aferida em metros.
Figura 6. Curva de crescimento e força de crescimento ontogênica.
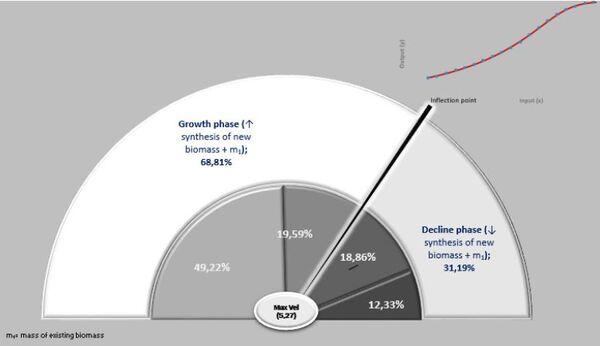
Primeiramente é necessário recordar a lei dos rendimentos decrescentes: há cada vez menor retorno adicional à medida em que adiciona-se uma mesma quantidade de recursos (Brue, 1993; Kuhi et al. 2012). Esse fato é observado na fase de declínio, a partir do ponto de inflexão. Assim, o primeiro período seguiria a lei dos “rendimentos crescentes”, defendida por Adam Smith (Vasconcellos, 1983).
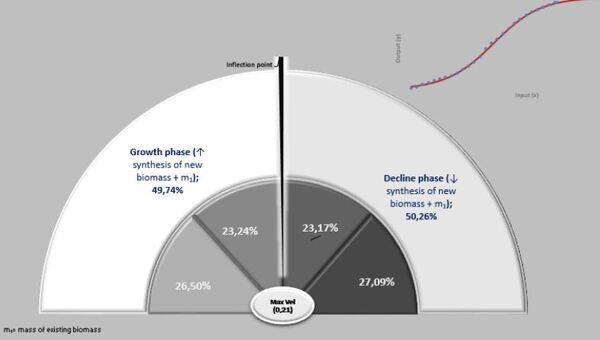
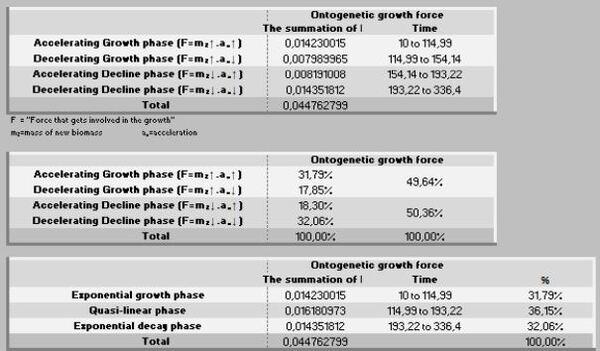
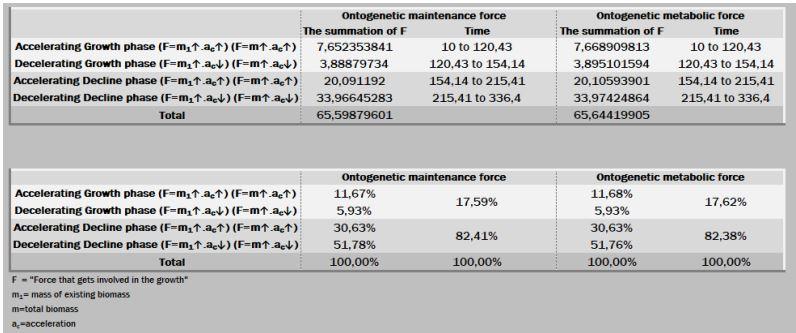
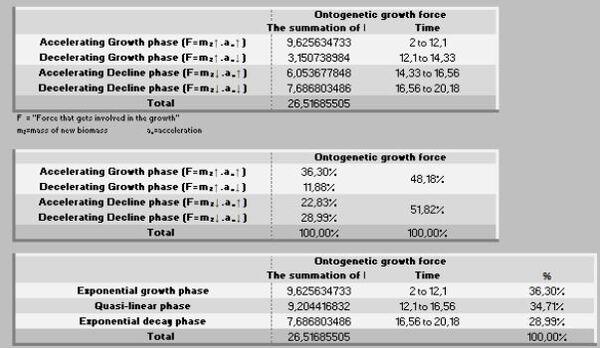
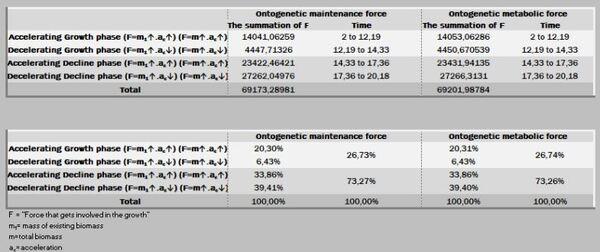
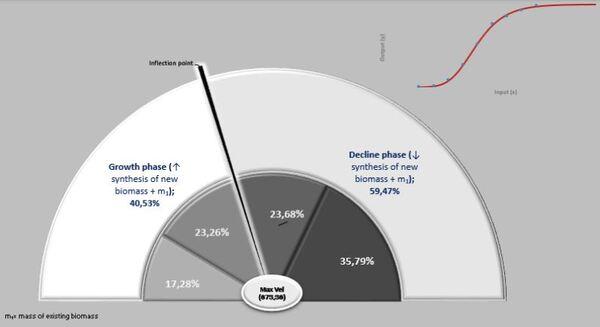
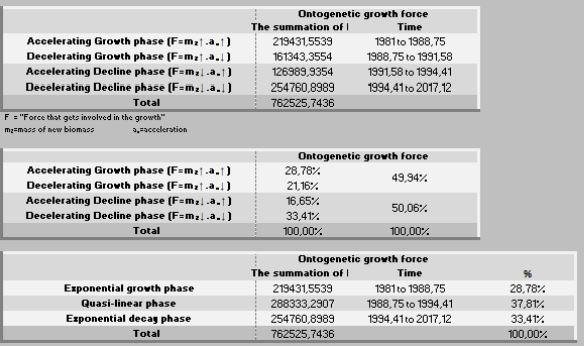
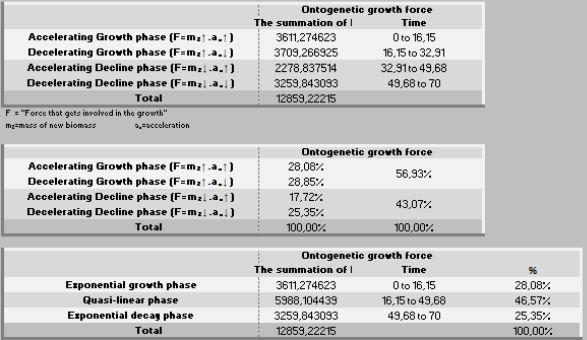
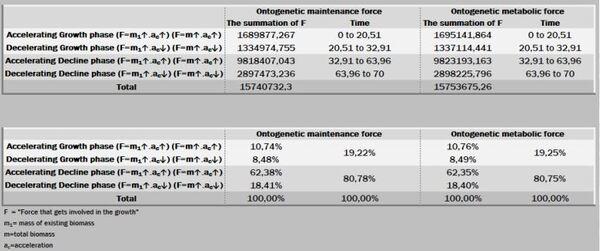
A ação das forças metabólicas na fase ascendente (lei dos rendimentos crescentes) se subdivide em dois momentos [ascendência acelerada (13,88%) e desacelerada (7,05%)], totalizando-se dos recursos energéticos de todo o crescimento. Decorrendo que 79,05% do recurso restante é subdividido em 36,39% no momento da fase de declínio em aceleração e 42,67% da desaceleração do declínio (Figura 7, 8 e 9).
Na prática, no período de 265 anos, nos primeiros 154 anos consomem 20,95% do total energético disponibilizado para o crescimento, sendo 79,05% para a fase dos rendimentos decrescentes. Ou seja, a manutenção é caríssima! Todavia, as forças ontogênicas de crescimento ficam bem balançadas em ambos os períodos, 50,62% no ascendente e 49,38% no declínio.
Provavelmente, o início da fase de saturação deve se dar quando as raízes atingem seu limite de extração de meios para crescer mais, e neste ponto todos os recursos são carreados somente para a manutenção da biomassa acumulada ao longo desses anos.
Figura 7. Fases de crescimento e declínio e forças atuantes.
Figura 8. Trajetórias das forças ontogênicas de crescimento, manutenção e metabólica.
Figura 9. Soma das forças ontogênicas de manutenção e metabólica.
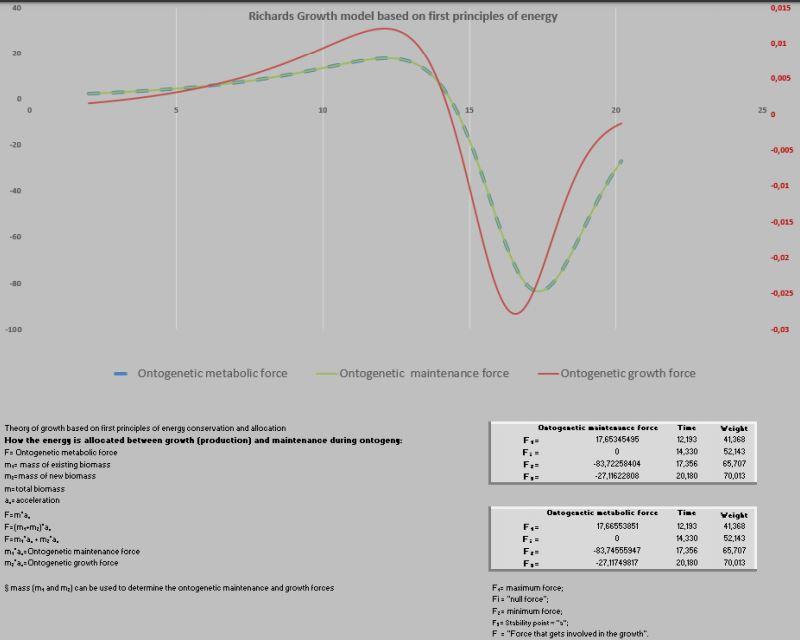
Exemplo 2 – Predição do peso de crianças
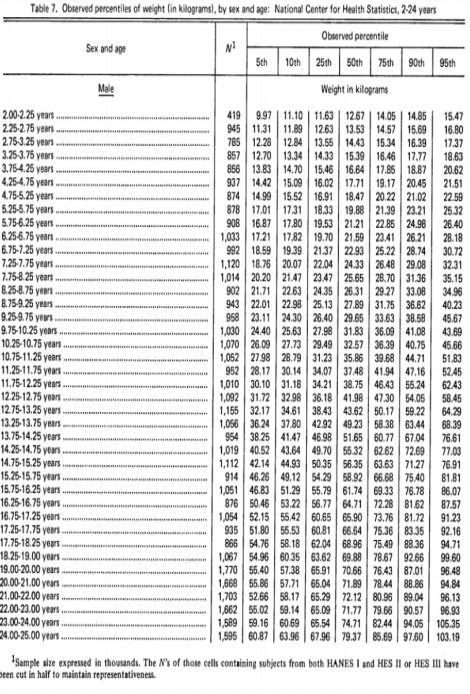
E no crescimento em peso de humanos (meninos) como seria a distribuição das forças metabólicas? (Figura 10)
Figura 10. Tabela americana de curvas de crescimento para crianças.
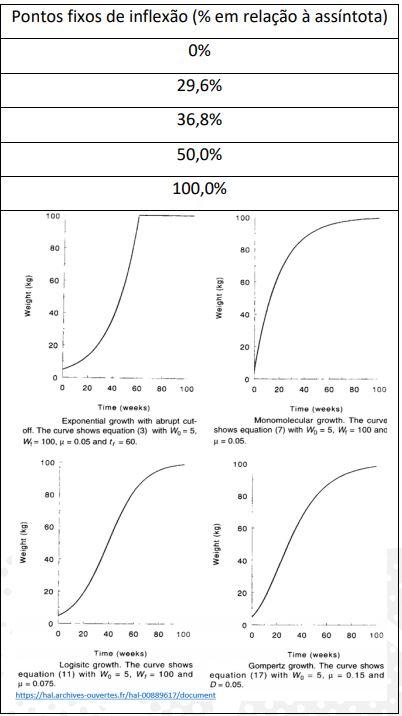
A curva é muito diferenciada, por apresentar um ponto de inflexão muito alto (73,71%; 52,14/70,74). Assim, os modelos de Von Bertanlaffy, Gompertz ou Logístico não seriam favoráveis para representar a curva de crescimento em humanos (Figura 11). Isso porque, Von Bertanlaffy, Gompertz e Logístico apresentam seus pontos de inflexão fixos em 29,63%; 36,79% e 50%, respectivamente de suas curvas (Goshu & Koya, 2013)
Figura 11. Curva ajustada pela planilha PPM e subsequente primeira e segunda derivadas (velocidade e aceleração).
A pergunta é: quando seria apropriado utilizar os modelos de Gompertz, Von Bertanlaffy, e Logístico? Apenas quando o pesquisador tiver certeza de que o ponto deinflexão é compatível com seus dados (Marinakis, 2012). Para piorar, o ponto de inflexão é influenciado pela genética, nutrição, manejo e ambiência (Knizetova et al. 1983, 1984; Aggrey, 2002).
Diante disso, por não ter como “garantir” o local do ponto de inflexão, o mais prudente seria escolher o modelo em que o ponto de inflexão não é fixo (ex: Weibull e Richards).
Assim sendo, retomando a curva para humanos (2 anos até 20 anos), pode-se observar que de 2 até 12,1 anos a criança tem seu período mais promissor de crescimento (fase de crescimento exponencial) e de 12,1 até 16,6 anos seria o crescimento quase-linear (F1 – F2). Dos 16,6 até 20,2 seria a fase exponencial de declínio e finalmente aos 20,2 anos a saturação do crescimento (Figura 12).
Figura 12. Curva de crescimento e força de crescimento ontogênica.
Do total de energia destinada para o crescimento, nesse período de 20 anos, 48,18% são consumidas até os 14,3 anos e 51,82% na fase de declínio. Por outro lado, do montanteda força destinada à manutenção utiliza-se 26,73% até os 14,3 anos e os restantes 73,27% no período de declínio. Coerentemente, a manutenção se mostra cada vez mais onerosa com o passar do tempo (Figuras 13 a 15).
Figura 13. Fases de crescimento e declínio e forças atuantes.
Figura 14. Trajetórias das forças ontogênicas de crescimento, manutenção e metabólica.
Figura 15. Soma das forças ontogênicas de manutenção e metabólica.
Exemplo 3 – Predição de casos de Aids
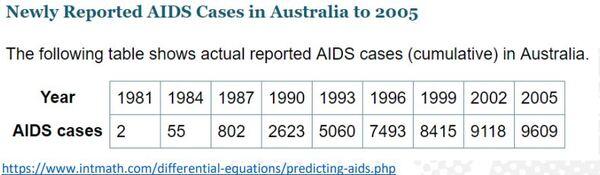
Nosso próximo exemplo será ajustar e prever casos de Aids até 2017, mas estando em 2005. O site sugere o modelo Logístico (ponto de inflexão fixo em 50%) (https://www.intmath.com/differential-equations/predicting-aids.php), porém com um ponto de inflexão livre obtém o valor de 40,54% (3960/9769). Valor esse acima de Von Bertanlaffy (29,63%) e Gompertz (36,79%) e abaixo da Logística (50%) (Figura 16).
Figura 16. Casos de Aids na Austrália (1981-2005).
Novamente, a melhor opção continua na escolha de modelos com o ponto de inflexão livre (Kuhi et al. 2010).
Agora é possível escolher até onde desejamos avançar para o futuro, alterando o “Stopping critical point”, que foi fixado em 99,9% no presente exemplo (Figura 17).
Figura 17. Curva ajustada pela planilha PPM e subsequente primeira e segunda derivadas (velocidade e aceleração).
Como a assíntota nunca atinge o ponto de máximo, há necessidade de limitar o seu máximo, sendo sugerido valor entre 90 a 99% (Passos et al., 2012).
Assim, cada caso é um caso!
Nossa recomendação é que prevalece o bom senso ao atender o objetivo da pesquisa
Nos casos de Aids na Austrália, conforme o modelo ajustado, o pico de contaminação ocorreu em 1991 (873 novos casos) de um total de 3960 pessoas contaminadas, decaindo a partir desse ano (Figuras 18 a 21).
Conforme os dados da planilha PPM, haverá uma acomodação em torno de 9769 casos. Esse montante ficará estabilizado em virtude dos novos casos, proporcionalmente, passarem a repor os óbitos, razão do valor da assíntota permanecer constante no referido valor.
A estatística não permite avaliar ou amenizar a dor que envolve os casos de Aids, entretanto pode orientar no entendimento da sua trajetória (R2 =99,924%).
O modelo de Richards mostra-se também muito oportuno para avaliar em tempo real surtos epidemiológicos, como o da Zika (Sebrango-Rodriguez et al., 2017), da dengue, H1N1 e SARS (Wang et al., 2012).
Figura 18. Curva de crescimento e força de crescimento ontogênica.
Figura 19. Fases de crescimento e declínio e forças atuantes.
Figura 20. Trajetórias das forças ontogênicas de crescimento, manutenção e metabólica.
Figura 21. Soma das forças ontogênicas de manutenção e metabólica.
Exemplo 4 – Frangos de corte
Adentrando em nosso projeto, os frangos de corte machos apresentam como limite máximo de crescimento o peso de 7,99 kg, contudo para isso, teoricamente, seriam necessários aproximadamente 218 dias para atingir esse peso (99,9% do valor “a”). Mas, na prática, segundo tabelas próprias para frango de corte, o limite máximo de alojamento estaria em 70 dias (Manual Ross 2017, Figura 22). Assim, como desejamos obter os pontos críticos F1, Fi, F2 e F3, e determinar as trajetórias do crescimento (fases) (Figura 23), optamos pela idade máxima de 70 dias (77,3% de “a”). Para isso, basta utilizar a barra de rolagem disponível de “99,9%” para “77,3%”.
Figura 22. Desempenho para frangos de corte machos (1-70 dias de idade).
Figura 23. Curva ajustada pela planilha PPM e subsequente primeira e segunda derivadas (velocidade e aceleração).
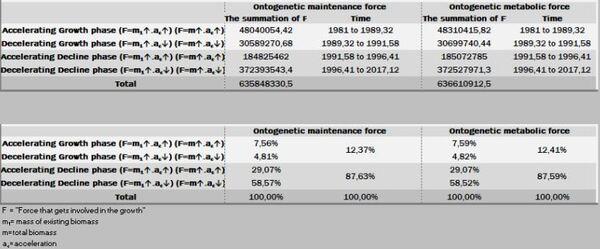
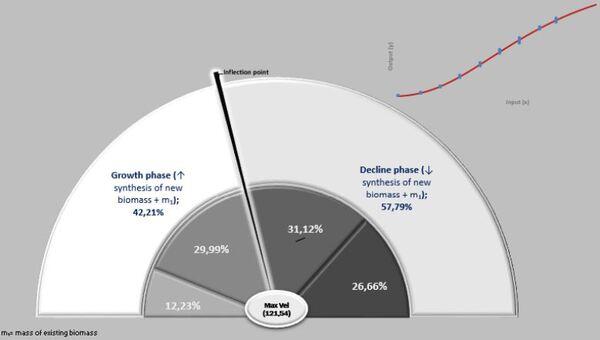
No presente experimento, os frangos machos foram capazes de ganhar 121,54 g/dia aos 32,9 dias de idade, decrescendo esse ganho a partir dessa data (Figuras 24 a 27).
Ao fixar o limiar de 70 dias para nossa trajetória de crescimento, o peso previsto para esse limite foi 6,177 kg. Isto é, 42,21% do seu ganho são obtidos em fase dos rendimentos crescentes (peso inicial até Fi) e 57,79% na fase dos rendimentos decrescentes (Fi – F3).
Figura 24. Curva de crescimento e força de crescimento ontogênica.
Figura 25. Fases de crescimento e declínio e forças atuantes.
Figura 26. Trajetórias das forças ontogênicas de crescimento, manutenção e metabólica.
Figura 27. Soma das forças ontogênicas de manutenção e metabólica.
A idade em F1 é 16,15 dias, segundo a força de crescimento ontogênica (Figura 24). A próxima fase, denominada em nossa planilha fase “Quase-linear” (F1 até F2) acomoda, também, a idade no ponto de inflexão da curva (≈ 33 dias).
Até o ponto F2 temos “todos os ventos” favoráveis ao crescimento da ave. Entretanto, esse cenário tranquilo muda na próxima fase que é definida como fase exponencial de declínio (F2 até F3). Esse seria um momento extremamente crítico, sendo bem explicado pela acentuada piora da conversão alimentar
Por fim, o ponto de saturação do crescimento (F3) no qual o trato digestivo atinge o “consumo máximo”, ou seja, nossa fonte energética. Isso inviabiliza manter qualquer ganho adicional desse ponto em diante, apenas mantém-se a massa que foi acumulada durante todo o período de crescimento. Vale lembrar que no início tinha apenas 45 g de massa (pintinho) e agora estamos com 5769 g em F2 e 6171 g em F3.
Do exposto, definir forças é uma área de pioneirismo (West at al., 2001), por adentrar nas “forças envolvidas na trajetória de crescimento” (Shimojo, 2006). Nessa nova proposta, a força ontogênica para manutenção envolve todos os esforços da ave para manter-se viva. No cotidiano por exemplo, significa andar, respirar, beber, comer, defender-se e por fim, crescer (força ontogênica do crescimento). Ou seja, a soma destas duas forças resulta na força metabólica ontogênica.
No período ascendente do crescimento (Peso inicial – Fi) há o investimento de 19,25% de recursos na manutenção da massa e, posteriormente no período de declínio, os restantes 80,75% dessa energia. Por isso, essa segunda fase é uma devoradora de recursos energéticos.
Por outro lado, a força ontogênica do crescimento, nesse específico intervalo (1-70 dias de idade) foi direcionada 56,93% para um primeiro período (peso inicial - Fi) e 43,07% para o segundo período (Fi-F3). Por sua vez, esses dois estágios podem ser desdobrados em: 1- fase exponencial de crescimento (28,08%), 2- fase quase-linear (46,57%), 3- na fase mais crítica do crescimento (exponencial decrescente), que consome 25,35% de energia.
Portanto, além do ponto de inflexão (Fi), é fundamental avaliar os pontos F1, F2 e F3 para plena avaliação de uma curva de modelagem.
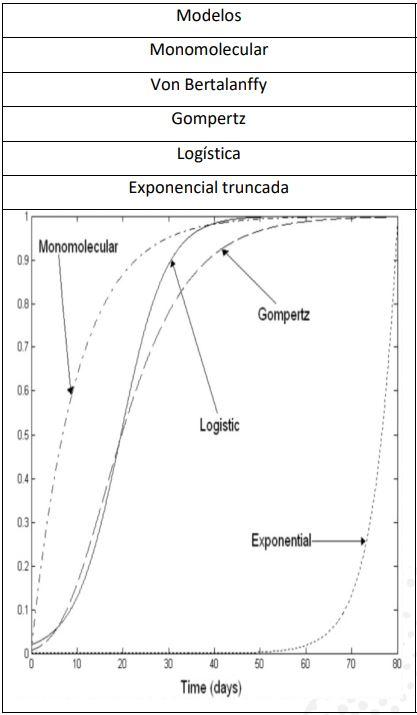
A curva de Richards pode apresentar seu ponto de inflexão desde o 0% (curva monomolecular) ou até o extremo de 100% (curva exponencial truncada) (Jorgensen,1981; France et al., 1996; Marinakis, 2012; Beiki et al., 2013; Selvaggi et al., 2015). No referido exemplo, temos o valor de 32,8% (2621/7991) para o ponto de inflexão, o qual foi intermediário entre os modelos de Von Bertanlanffy e Gompertz (Figura 28).
Figura 28. Pontos fixos de inflexão de curvas sigmoides.
Conclui-se que o modelo de Richards, por apresentar a habilidade de ter seu ponto em qualquer local da curva, mostra-se mais adequado para interpretações biológicas,podendo ainda acomodar os modelos anteriormente citados (Família Richards) (Knizetová, et al., 1984; Kuahi et al., 2010; Teleken et al., 2017).
Além disso, a avaliação da ontogenia (do nascimento à maturidade) das forças de crescimento poderá expressar melhor os aprimoramentos genéticos, nutricionais, de manejo e ambientais em diversos ramos biológicos para obter a máxima eficiência econômica.
Do ponto de vista energético, quanto mais próximo o abate estiver do ponto Fi, mais eficiente será, pois o recomendado é que realize o abate entre Fi e F2, mas nunca depois de F2 (Knizetová et al. 1995).
Em resumo, há de se tomar muito cuidado com o modelo escolhido para avaliar a trajetória de crescimento de um evento, pois poderemos apenas obter soluções matematicamente corretas, porém biologicamente inviáveis (Fitzhugh 1976, Harvey & Ransnas, 1987, Brown, 2001; Porter et al. 2010; Kemmer & Keller, 2010; Korkmaz & Uckardes, 2013; Firat et al. 2016).
Certamente, o objetivo primordial é descrever a sequência: dimensão versus tempo, em poucos parâmetros e predizer taxas de crescimento, requisitos nutricionais, respostas de seleção genética e outros itens de interesse (Fitzhugh 1976; Eleroglu et al, 2014).
Discussão geral
A eficiência técnica permite através do melhoramento genético, manejo e nutrição uma melhor conversão alimentar para produzir a mesma quantidade de kg de frangos (utilização de menor quantidade de nutrientes). Outra visão seria almejar a eficiência econômica, na qual teríamos como meta a mesma produção em kg de frangos, mas com o menor custo possível (Nahashon et al., 2006; Selvaggi et al., 2015). Assim, independente do ponto de vista, não se sustenta uma produção viável de frangos de corte sem rendimentos.
É definido pela curva sigmoide, e principalmente pela de Richards que apresenta o ponto de inflexão móvel, três distintos períodos: 1- Fase de Retornos Crescentes (Lei dos Rendimentos Crescentes); 2- Fase Quase-Linear (Constante de Escala); 3- Fase de Retornos Decrescentes (Lei dos Rendimentos Decrescentes).
Diante disso, os investimentos realizados durante todo o ciclo de produção do nosso moderno frango de corte (Nutrição, Genética e Sanidade e Ambiência) podem ser perfeitamente observados em sua curva de crescimento ontogênica, que vai desde o nascimento até sua maturidade, salientando que o abate deveria ocorrer antes do ponto F2 (Makarieva et al., 2004; Rocha-Silva et al., 2016).
É indispensável reforçar o já abordado: a curva sigmoide por si só, apesar de muito estudada, não aflora todo seu potencial apenas em seus tradicionais três ou quatro parâmetros; portanto, faz-se necessário adentrar em suas derivadas primeira e segunda para garimpar mais riquezas. Várias pesquisas já se dedicam a isso (Gregorczyk, 1991; Shimojo et al. 2002; Berzsenyi & Lap, 2004; Shimojo, 2006; Scapim & Bassanezi, 2008; Shimojo et al. 2008, 2012; Shimojo,2016; Swain et al. 2016). Por incrível que pareça, a curva sigmoide ainda esconde muito mais recursos (Shimojo et al. 2009, 2010).
Com certeza, bastava “cavar” um pouco mais para encontrar o maior tesouro escondido pela curva sigmoide (Figura 29). Ainda é necessário relembrar que essa preciosidade foi apresentada por Shimojo et al. (2006) quando propõe F= m. a (Figuras 30 a 32).
Figura 29. Necessitamos cavar um pouco mais!
Figura 30. Curva de crescimento da massa existente e nova biomassa de frangos de corte.
Esse “óbvio” que ninguém enxergava, até que Shimojo (2006) expressou com simplicidade. Utilizando-se dessa informação, a planilha PPM viabiliza multiplicar a massa de cada dia vezes a aceleração desse mesmo período para obter a força nesse ponto (Força Ontogênica metabólica).
Figura 31. Curva da velocidade (primeira derivada).
Desse ponto de vista mecanístico oferecido por Shimojo et al. (2006), podemos ainda deduzir que esse “peso total” se subdivide em massa já existente (m1= foto de frango ) e a nova biomassa (m2 = foto de frango )
Figura 32. Curva da aceleração (segunda derivada).Figura 32. Curva da aceleração (segunda derivada).
Outros trabalhos, muito próximo desta proposta, são relatados por Hozumi (1989), Von-Stockar & Liu (1999), West et al. (2001, 2002), Urban (2002), West et al. (2004), Morowitz & Smith (2007), Enberg et al. (2008), Hook et al., 2011, Maino & Kearney (2015).
Novamente, o óbvio invisível (via-se, mas sem enxergar) aflora. A curva (força) correta para definir os pontos F1, F2 e F3 na curva de crescimento, obrigatoriamente, é a força “Ontogênica de Crescimento”.
Efetivamente, as três fases do crescimento de uma curva sigmoide são:
1- Fase de Crescimento Exponencial (Peso ao Nascimento – F1);
2- Fase Quase-Linear (F1 – F2);
3- Fase de Decréscimo Exponencial (F2 – F3).
Sendo F3 o peso de maturidade do animal.
Podemos assegurar que a grande beleza da curva sigmoide está nessa “força de crescimento”, definida pelo produto da segunda derivada pela nova biomassa, que permite determinar os pontos F1, Fi, F2 e F3.
Esses importantes pontos (F1, Fi, F2 e F3) serão muito úteis para avaliar não só o progresso genético das empresas melhoradoras, como também o aprimoramento em nutrição ao comparar níveis e novos produtos. Além disso, apreciar novas propostas de manejo e também de sanidade e ambiência.
Talvez estamos muito aquém do entendimento do potencial e aplicação das curvas sigmoides. A literatura mais aplicada e prática parece ser a empresarial. Nessa abordagem, o ponto mais importante seria definir o momento da transição (Ponto A). Esse ponto, se ignorado, mostra-se fatal. Foi o caso das empresas Kodak e Britânica (https://www.youtube.com/watch?v=iJmvTJ4VSxo). Assim sendo, temos muito que aprender com as empresas e suas similaridades em nossa área (Arantes, 2012).
Por exemplo, ao considerar o ponto de abate qual o foco mais favorável: eficiência técnica ou econômica? Essa resposta está na curva sigmoide! E cada caso será um caso. Ou seja, teremos que rodar em cada novo ciclo uma nova curva (no tempo presente, olhando o passado, para prever o futuro) para avaliar as forças favoráveis e desfavoráveis resultantes das interações de todos os fatores envolvidos no processo (Quadro 1).
Portanto, fica a questão: “Onde você está na curva”?
Quadro 1. Curva sigmoide e suas aplicações práticas.
Assim, para qualquer processo de crescimento haverá uma curva para apresentar o acumulado no tempo. O conhecimento é algo que cresce com o tempo. O novo aprendizado seria nossa primeira derivada (velocidade que acumulamos novos conhecimentos) e a taxa de progresso (segunda derivada) que avalia a aceleração do aprendizado. Conforme o provérbio de Salomão (Pv 19:27): “Se você parar de aprender, logo esquecerá o que sabe”.
O perigo continua sendo o ponto “A” (nosso ponto “F2”), a partir do qual o acúmulo de novos conhecimentos tende a cair exponencialmente. Em consequência disso, o profissional pode tornar-se desatualizado e fatalmente desqualificado para enfrentar um mercado cada vez mais exigente.
Em suma:
A proposta de “dissecar” as equações das curvas sigmoides já vem de longa data, valendo-se do uso de ajustes à mão livre, com posterior linearização da equação e com a sobreposição de “transparências” com curvas padrões. Finalmente o cálculo dos parâmetros por regressão (Barusso & Ribeiro, 1976).
O problema de linearização está na perda da originalidade dos dados e armadilhas nas análises que podem levar a erros nas conclusões (Brisbin et al., 1987; Brown, 2001; Venugopalan & Shamasundaran, 2003).
Devido aos avanços na informática e atualmente com auxílio de planilhas foi possível grande avanço no estudo das curvas de crescimento e, principalmente, na tentativa de atualizar seus parâmetros de forma biológica (Yin et al.,2003).
Assim, os parâmetros foram avaliados e revirados para extrair desses o máximo de informações (Knizetová et al. 1983, 1984, 1991; Aggrey 2002; Sengul & Kiraz, 2005; Cetin et al., 2007; Eleroglu et al., 2014).
Outras estratégias passaram a ser utilizadas como a primeira e a segunda derivadas da curva de crescimento, objetivando determinar os componentes da velocidade e a aceleração (Nath & Moore, 1992; Gregorczyk, 1998; Berzseny & Lap, 2004), tendo em vista os pontos P1, Pi e P2, como reforços aos parâmetros tradicionais.
A saga continua ao se aprofundar no misterioso campo do anabolismo e catabolismo embutidos nas curvas sigmoides (Hozumi, 1989).
Entretanto, relembrando que o “ovo de Colombo” foi oferecido por Shimojo et al. (2006) ao introduzir o ponto de vista mecanístico (Leis de Newton) na análise das curvas de crescimento. O óbvio, porém difícil de enxergar, até que Shimojo et al. (2006) o expressa com simplicidade F= m.a !
Tão óbvio e tão simples e agora aplicado na planilha PPM.

















.jpg&w=3840&q=75)