Curvas de crescimento para codornas de corte
1Programa de Pós-graduação em Zootecnia (PPGZOO), Universidade Federal dos Vales do Jequitinhonha e Mucuri (UFVJM), 2Mestre em Zootecnia, PPGZOO, UFVJM, Diamantina, MG, Brasil, 3Curso de Zootecnia, Departamento de Zootecnia (DZO), UFVJM, Diamantina, MG, Brasil, 4DZO, UFVJM, Diamantina, MG, Brasil
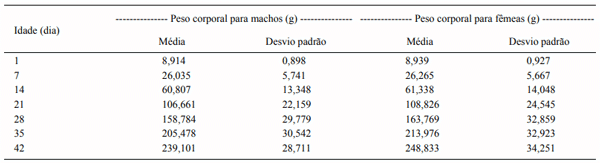
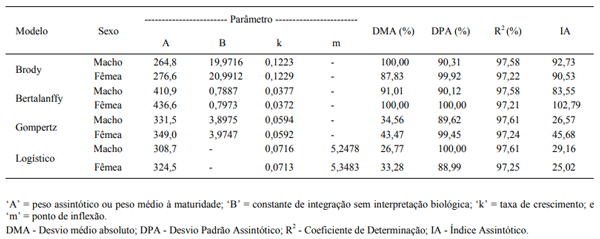
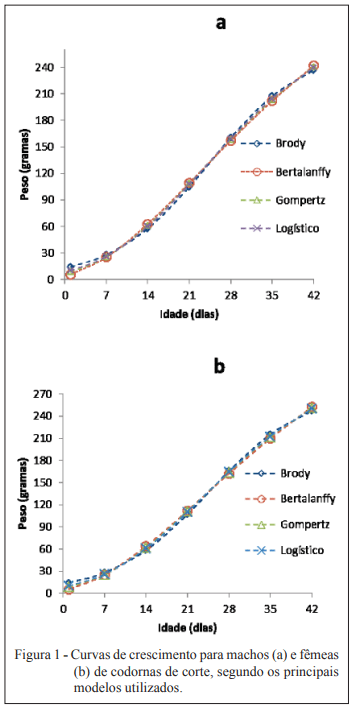
Objetivou-se com este trabalho comparar modelos não lineares para descrever o crescimento de codornas de corte machos e fêmeas do nascimento ao 42o dia de idade. Foram utilizados 24.679 registros de peso corporal de codornas de corte da linha fêmea LF1, pertencente ao Programa de Melhoramento de Codornas da UFVJM, em Diamantina, MG. As codornas de três gerações sucessivas foram pesadas a cada sete dias (nascimento, 7, 14, 21, 35 e 42 dias de idade). Os modelos de Brody, von Bertalanffy, Richards, Logístico e Gompertz foram ajustados ao conjunto de dados observados. Os parâmetros dos modelos foram estimados, para machos e fêmeas separadamente, pelo algoritmo de Gauss Newton, utilizando-se o procedimento NLIN do SAS. Os critérios utilizados para escolha do modelo de melhor ajuste da curva de crescimento foram o coefi ciente de determinação (R2 ), o desvio padrão assintótico (DPA), o desvio médio absoluto dos resíduos (DMA) e o índice assintótico (IA). Dos cinco modelos utilizados, apenas o modelo de Richards não apresentou convergência, tanto para machos como para fêmeas. O menor valor para o índice assintótico (IA) foi observado para o modelo de Gompertz em machos (26,57) e o Logístico para fêmeas (25,02), indicando serem estes os modelos que mais se ajustaram aos dados.
Palavras-chave: Coturnix coturnix coturnix, curva de crescimento, modelos não lineares.

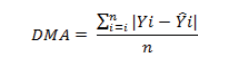

ARAÚJO, R.O. Componentes de covariâncias estimados por metodologia bayesiana para parâmetros biológicos obtidos por modelos não lineares para bubalinos da raça Murrah. 2009. 73f. Dissertação (Mestrado em Produção Animal) - Programa de Pósgraduação em Zootecnia, Universidade Federal de Santa Maria, RS.
BERTALANFFY, L. Quantitative laws in metabolism and growth. Quarterly Review of Biology, v.32, n.3, p.217-230, 1957. Disponível em: . Acesso em: 14 nov. 2011.
BRODY, S. Bioenergetics and growth. New York: Reinhold, 1945. 1023p. Disponível em: . Acesso em: 16 nov. 2011. BROW, J.E. et al. A comparison of nonlinear models for describing weight-age relationships in cattle. Journal of Animal Science, v.42, n.4, p.810-818, 1976. Disponível em: . Acesso em: 22 nov. 2011.
CORRÊA, G.S.S. et al. Exigências em proteína bruta para codornas de corte EV1 em crescimento. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.59, p.1278-1286, 2007. Disponível em: . Acesso em: 23 nov. 2011. doi: 10.1590/S0102-09352007000500027.
FREITAS, A.R. et al. Modelos de curva de crescimento em frangos de corte. Pesquisa Agropecuária Brasileira, v.19, n.9, p.1057- 1064, 1984. Disponível em: . Acesso em: 02 de dez. 2011.
GARNERO, A. del V. et al. Parâmetros genéticos da taxa de maturação e do peso assintótico de fêmeas da raça Nelore. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.57, n.5, p.652-662, 2005. Disponível em: . Acesso em: 25 nov. 2011. doi: 10.1590/S0102-09352005000500011.
GOUS, R.M. Lysine: making progress in the nutrition of broilers. Poultry Science, v.77, n.1, p.111-117, 1998. Disponível em: . Acesso em: 14 nov. 2011.
HARTLEY, H.O. The modifi ed Gauss-Newton method for the fi tting of nonlinear regression functions by least squares. Thechnometrics, v.3, p.269-280, 1961. Disponível em: . Acesso em: 16 nov. 2011.
KNIZETOVA, H. et al. Analysis of growth curves of fowl. I - Chickens. Poultry Science, v.32, p.1027-1038, 1991. Disponível em: . Acesso em: 16 nov. 2011. doi: 10.1080/00071669108417427.
LAIRD, A.K. Dynamics of relative growyh. Growth, v.29, p.249-263, 1965. Disponível em: . Acesso em: 18 nov. 2011.
LAIRD, A.K.; HOWARD, A. Growth curves in imbred mice. Nature, v.213, n.5078, p.786-788, 1967. Disponível em: . Acesso em: 13 nov. 2011. doi: 10.1038/213786a0.
MANSOUR, H. et al. Analysis of covariance structure of repeated measurements in holstein conformation traits. Journal of Dairy Science, v.74, n.8, p.2757-2766, 1991. Disponível em: . Acesso em: 15 nov. 2011.
MAZUCHELI, J. et al. Modelo de crescimento de Gompertz na presença de erros normais heterocedásticos: um estudo de caso. Revista Brasileira de Biometria, v.29, n.1, p.91-101, 2011.
NELDER, J.A. The fi tting of a generalization of the logistic curve. Biometrics, v.17, n.1, p.89-110, 1961. Disponível em: . Acesso em: 11 nov. 2011.
OLIVEIRA, E.G. et al. Desempenho produtivo de codornas de ambos os sexos para corte alimentadas com dietas com quatro níveis protéicos. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.7, n.2, p.75-80, 2002. Disponível em: . Acesso em: 19 nov. 2011.
RAJINI, R.A.; NARAHARI, D. Dietary energy and protein requeriments od growing Japanese quails in the tropics. Indian Journal of Animal Sciences, v.68, n.10, p.1082-1086, 1998.
RATKOWSKY, D.A. Handbook of nonlinear regression models. New York and Basel, Marcel Dekker, 1990. 241p.
RONDON, E.O.O. et al. Modelagem computacional para produção e pesquisa em avicultura. Revista Brasileira de Ciência Avícola, v.4, n.1, p. 199-207, 2002. Disponível em: . Acesso em: 22: nov. 2011.
SARMENTO, J.L.R. et al. Estudo da curva de crescimento de ovinos Santa Inês. Revista Brasileira de Zootecnia, v.35, p.435- 442, 2006. Disponível em: . Acesso em: 27 nov. 2011.
SAS Institute. SAS/STAT® user’s guide: version 6. 4.ed. Cary, NC, 1990. p.1022.
SILVA, F.F. et al. Infl uência de fatores genéticos e ambientais sobre as estimativas dos parâmetros das funções de crescimento em gado nelore. Ciência Agrotécnica, v.25, n.5, p.1195-1205, 2001. Disponível em: . Acesso em: 27 nov. 2011.
THOLON, P.; QUEIROZ, S.A. Modelos matemáticos utilizados para descrever curvas de crescimento em aves aplicados ao melhoramento genético animal. Ciência Rural, v.39, n.7, p.2261- 2269, 2009.
VELOSO, R.C. et al. Níveis de proteína bruta e energia metabolizável para codornas de corte. Acta Scientiarum Animal Sciences, no prelo, 2012.







