INTRODUÇÃO
Os maiores componentes das dietas de suínos em vários países são milho, soja, cevada e/ou trigo (PEET-SCHEWERING et al., 1991). O crescimento da população mundial provoca aumento da competição entre animais não-ruminantes e pessoas por grãos de alta densidade nutritiva (JOHNSTON et al., 2003). Com isso, o futuro da alimentação de espécies animais que utilizam fundamentalmente grãos de cereais, como é o caso dos suínos, deve ser reavaliada e atenção especial deve ser dada à habilidade desses animais em utilizarem alimentos alternativos como fonte de energia, não utilizável pelo homem e de baixo custo para o produtor (TEIXEIRA, 1995).
Palhada e outros subprodutos são inevitavelmente produzidos durante a produção de cereais. Sabese, também, que no futuro, cereais e outros produtos serão necessários para o consumo humano. Resíduos da agricultura são tradicionalmente utilizados para muitos propósitos, dentre eles a alimentação de animais ( FAO, 1977).
O custo da alimentação representa mais de 60% do total da produção de suíno e a energia é o maior componente (NOBLET e PEREZ, 1993). Diante desse quadro, o efeito de dietas fibrosas sobre a fisiologia digestiva dos animais está gerando cada vez mais interesse, principalmente entre os monogástricos, onde o conhecimento de microorganismos envolvidos na quebra da fibra é ainda limitado, quando comparado com animais poligástricos (FONTY e GOUET, 1989).
O suíno é considerado como animal não-ruminante de ceco não-funcional, ao contrário dos eqüinos, por exemplo, considerados de ceco funcional (CLOSE, 1994). Os componentes dietéticos da fibra são minimamente digeridos no intestino delgado de suínos, providenciando substrato para a fermentação microbiana no intestino grosso. O principal produto desta fermentação são os ácidos graxos voláteis (AGV) propionato, butirato e acetato. A contribuição calórica destes AGV em suínos tem sido estimada em 5% a 28% das exigências em energia de manutenção, dependendo da freqüência do consumo e do nível de fibra na dieta (NRC, 1998).
O suíno, ainda, é um dos melhores modelos para o homem quanto a digestão. É um animal onívoro monogástrico, cujo trato digestivo posterior é bem desenvolvido devido ao seu hábito alimentar (FONTY e GOUET, 1989).
Porém, a presença de dieta fibrosa reduz significativamente a digestibilidade de todos os nutrientes; há uma redução de consumo de energia, influenciando numa queda do crescimento e da eficiência da conversão alimentar, mas proporcionando uma carcaça mais magra (CLOSE, 1994).
A possibilidade de se aumentar o uso de subprodutos de elevado teor fibroso e forragens na produção de suínos tem sido estudado por diversos pesquisadores e os resultados tem demonstrado que as porcas são mais aptas para utilizar fibra do que suínos desmamados ou em fase de crescimento/ terminação . Estas apresentarão melhoras reprodutivas, assim, como na saúde e no bem estar (CLOSE, 1994). Os suínos jovens, devido ao menor desenvolvimento do trato gastrointestinal, tem menor área para a disseminação da população microbiana resultando num menor aproveitamento do alimento fibroso (TEIXEIRA, 1995), embora WOJCIK et al. (1991) utilizando a raça Large-White, observou resultados com ganho de até 613g dia-1 com suínos na fase de crescimento/terminação quando receberam dietas de até 11,85% de fibra.
Ressalta-se, ainda, que há uma considerável capacidade de adaptação da morfologia e da flora do trato gastrointestinal desses animais às dietas fibrosas, sendo de grande importância para a intensidade de degradação da fibra a sua composição química e origem (TEIXEIRA, 1995).
Há, portanto, um potencial considerável para a incorporação de fibra e subprodutos em estratégias alimentares para suínos nos países em desenvolvimento, com economia e vantagem produtiva (CLOSE, 1994).
Constituintes da fibra
A) Celulose: A mais abundante macromolécula "in natura", é um polímero linear com ligações beta- 1-4 de milhares de unidades de glicose. Ocorre, principalmente, na forma cristalizada, organizada como microfibrilas, onde a cadeia de celulose é hermeticamente vedada junto à agregados compactos envoltos pela matriz de outros constituintes da parede celular. A cadeia de glucano é mantida unida por pontes de hidrogênio e entre cadeias adjacentes. Essa conformação favorece a formação de tais ligações e explica a força mecânica da celulose bem como sua resistência tanto à degradação enzimática e microbiana como a hidrólise ácida (THEANDER et al., 1989).
B) Hemicelulose: O termo hemicelulose foi, originalmente aplicado à polissacarídeos, os quais eram estruturalmente relacionados e associados e que eram preferencialmente solúveis em solução alcalina aquosa, após a remoção de polissacarídeos solúveis em água e polissacarídeos pectínicos, xilano e xiloglicano (componentes importantes da parede celular primária) pertencentes a este grupo na família de fibra dietética como outro tipo de polissacarídeos podem também necessitar condições alcalinas para serem extraídos no mesmo material (THEANDER et al., 1989).
C) Substâncias pectínicas: Refere-se à uma complexa mistura polissacarídica coloidal, a qual pode ser parcialmente extraída com água. Uma visão geral é que as pectinas são parcialmente esterificadas com metil ramogalacturonas, na qual unidades de raminose são interligadas, e que outros açucares como arabinose e galactose ocorrem em pequenas cadeias laterais (THEANDER et al., 1989). Os cereais se caracterizam por terem quantidades insignificantes de substâncias pectínicas. O endosperma da parede celular do trigo tem predominantemente, arabinoxilano como polissacarídeo, parcialmente solúvel em água, enquanto que na cevada e a aveia predomina 1,4-; 1,3-beta-glucano, respectivamente. Na camada aleuronica de trigo e cevada, predomina arabinoxilano e mistura de beta-glucano ligados (THEANDER et al., 1989).
D) Lignina: Pode ser descrita por uma rede ramificada de unidades fenilpropano nas fibras para aumentar sua rigidez. Acredita-se que os precursores desses compostos sejam os álcoois anamílicos, coniferílico e para-cumarílico os quais são transformados em lignina nas plantas através de um complexo processo de polimerização desidrogenativa (THEANDER et al., 1989).
Segundo VAN SOEST(1981) a composição química da lignina varia conforme a espécie da planta, a sua maturidade e seu desenvolvimento.
Tal afirmação é suportada por THEANDER et al. (1989) os quais relataram que a proporção das unidades à base de guacil e p- hidroxifenil formado em diferentes tipos de lignina é um detalhe fundamental, que diferenciará entre as plantas.
A lignina é parcialmente ligada à parede celular composta de hemicelulose e representa tipicamente a parte hidrofóbica do complexo da fibra bruta e irá contribuir nas propriedades físicas e nutricionais do intestino, por exemplo, ligando os ácidos biliares (THEANDER et al., 1989).
E) Sílica: Ocorre na maioria das plantas e pode ser uma parte estrutural da planta ou o resultado de uma contaminação de solo (VAN SOEST, 1981).
É encontrado com abundância em casca de arroz, palha de arroz e em muitas gramas tropicais (VAN SOEST, 1970) e parece reduzir a digestibilidade da matéria seca em 1 à 3 vezes (VAN SOEST e JONES, 1968; SMITH et al., 1971).
A remoção da sílica com álcalis aumenta a digestibilidade (VAN SOEST, 1970). A sílica total pode ser estimada como resíduo mineral da fibra detergente ácido insolúvel em hidróxido de bromo. A sílica do solo pode ser determinada se esse procedimento for precedido por extração fibra detergente neutro (VAN SOEST, 1981).
F) Cutina: A cutina é geralmente encontrado na cutícula (epiderme) das plantas. Nas cascas de arroz e amendoim o teor pode ser de 10 à 20% (VAN SOEST, 1969). Como a cutina é indigerível há uma redução na digestibilidade dos carboidratos da parede celular. A cutina pode ser avaliada utilizando sistema de análise de detergente, como o resíduo orgânico resistente à 72% de ácido sulfúrico e óxido de permanganato.
G) Ácidos fenólicos e grupos acetil: Ácidos fenólicos (ferrúlico, diferrúlico e para-cumálico) constituem 0,52 à 5% da parede celular em gramíneas (WAITE e CORROD, 1959; THEANDER et al., 1981), mas pode ser removido por álcalis sob condições relativamente brandas. Admite-se que ambos são unidos á hemicelulose (HARTLEY e JONES, 1977).
H) Taninos: Taninos ocorrem em muitas plantas e alguns tem capacidade por irreversibilidade ligar proteínas e inibir a fermentação da fibra. Eles interferem, também, na análise da fibra (VAN SOEST, 1981). A extração química de tanino, mostrou melhorar a digestibilidade da fibra. Nenhuma análise quantitativa tem sido até agora desenvolvida para cálculo do tanino especificamente interferindo com a digestão de fibra e /ou proteína (ROBBINS, 1973).
Conceito de fibra
Os polissacarídeos estruturais que constituem a parede celular dos vegetais são polímeros de pentose (arabinose e xilose) e hexoses (glicose, frutose e galactose). Estas unidades básicas combinam-se originando dois grupos estruturais principais: homopolissacarídeos e heteropolissacarídeos. No primeiro encontra-se a celulose e no segundo, as pectinas e as hemiceluloses. Estes dois grupos, formam a fração insolúvel ou carboidratos insolúveis, também chamados de polissacarídeos não amiláceos. Estes carboidratos, juntamente com a lignina (polímero aromático de ésteres fenólicos) e certos polissacarídeos de reserva, como as gomas, constituem a "fibra" (TEIXEIRA, 1995).
A fibra alimentar pode ser considerada como a fração do alimento não digerível pelas enzimas endógenas dos animais, mas sim pela ação de enzimas da microbiota gastrintestinal (TUNGLAND e MEYER, 2002).
Verifica-se, entretanto, grande variação na composição química da fibra de diferentes origens (TEIXEIRA, 1995). Portanto, dependendo do teor relativo da celulose, hemicelulose, pectina e lignina, a fibra bruta representa quase sempre apenas uma parte do real consumo de fibra do animal. Assim, dependendo da origem e, portanto, da composição química da fibra usada, mensurações baseadas na fibra bruta poderá induzir à estimativas inexatas da digestibilidade aparente da fibra da dieta (STANOGIAS e PEARCE, 1985).
Informações errôneas poderão, também, ser obtidas quando celulose purificada é utilizada como fonte de fibra da dieta, pois esta difere, consideravelmente, do material celulósico natural em suas propriedades físicas e biológicas e parece ser mais resistente do que a celulose natural diante da digestão microbiana no intestino grosso de suínos (STANOGIAS e PEARCE, 1985).
Assim, a precisão na análise da composição química da fração fibrosa do alimento ou da dieta deve ser a mais acurada possível para uma estimativa correta das porções degradáveis nos diferentes sítios do trato gastrointestinal (TEIXEIRA, 1995).
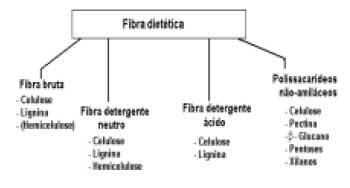
O tradicional "Método de Weende", utilizado para a determinação da fibra mostra-se impreciso, determinando a fibra bruta e os extratos nãonitrogenados, havendo apenas uma recuperação de cerca de 50% da celulose, 15% da hemicelulose e 10- 50% da lignina, ficando a porção de extrato nãonitrogenado com todos os erros acumulado no decorrer das análises (TEIXEIRA, 1995). Devido principalmente, à grande variação dos componentes químicos da parede celular, outros métodos foram desenvolvidos, como o uso de detergentes. O "Método de Van Soest", citado por SILVA (1990) determina a fração correspondente à parede celular vegetal, que é composta por celulose, hemicelulose, lignina e proteínas lignificadas (Fibra em Detergente Neutro – FDN) e a fração correspondente à celulose + lignina (Fibra em Detergente Ácido – FDA) (Figura 1).
Figura 1. Frações e componentes da fibra dietética.
Algumas técnicas modernas de maior exatidão tem sido desenvolvidas, onde a quantificação dos carboidratos da fibra é realizado utilizando cromatografia líquida, gasosa ou colorimétrica combinada com a determinação gravimétrica da lignina (THEANDER et al., 1989).
O melhor entendimento dos complexos efeitos do processo de assimilação de fibras pelos suínos só será possível a partir de um conhecimento mais detalhado da composição e das propriedades físico- químico das fibras presente na dieta.
Digestibilidade da fibra pelos suínos: Tipo de fibra
Suínos que receberam fibra de fonte purificada e fonte natural apresentaram uma digestão aparente variando de 21% à 92%, podendo concluir que a fonte de fibra é um fator importante na determinação da digestibilidade (STANOGIAS e PEARCE, 1985).
Segundo THEANDERet al. (1989) os farelos são particularmente mais ricos em fibra. As análises indicaram maiores teores de lignina, são mais ricos em arabinoxilano e celulose, mas tem um baixo teor de constituintes ácido urônico.
As cascas dos cereais são, na maior parte, compostas de componentes como arabinoxilano, celulose e lignina, enquanto que beta-glucano e quantidades variáveis de arabinoxilano estão presentes na fração do endosperma (THEANDER et al., 1989).
A melhor digestibilidade observada para suínos, da polpa da beterraba e da casca de soja quando comparada a outros alimentos que contem elevado teor de fibra, deve-se ao alto conteúdo de substâncias pécticas contidas nesses alimentos, sendo essas substância altamente digestíveis. Além disso, há um baixo grau de lignificação e baixa proporção de amido. Tais características diferenciam esses alimentos em termos de digestibilidades das demais matéria- prima ricas em fibra e de interesse para a alimentação de suínos (TEIXEIRA, 1995).
A propriedade higroscópica das fibras vegetais constitui um dos aspectos relevantes para explicar a digestibilidade dos alimentos. As substâncias pécticas estão, entre os polissacarídeos da parede celular, entre as que apresentam características mais acentuadas no processo de retenção de água. Esta propriedade reveste-se de importância à medida que proporciona aumento do tempo de exposição da digesta à ação enzimática e microbiana, embora segundo SAUER e OZIMEK (1986) a pectina promove a formação de uma substância, espécie de gel, que impede o acesso das enzimas digestivas no intestino delgado, passando a ser fermentado, então no intestino grosso.
BRIOLLOUET e CARRE (1983) relatam que as sementes leguminosas são ricas em substâncias pécticas, e que chegam a representar mais de 70 % dos componentes totais da parede celular.
HARTOG et al.. (1988) observaram que o coeficiente de digestibilidade ileal da proteína da dieta contendo palha de trigo foi significativamente inferior ao da dieta controle, enquanto o mesmo mostrouse semelhante entre as dietas contendo pectina, celulose e a dieta controle.
A inclusão de polpa de citrus (10 à 40%) em dietas para suínos em crescimento mostrou um aumento significativo da digestibilidade da fibra bruta e da celulose, indicando que os carboidratos contidos na polpa de citrus são altamente digestíveis por suínos (BAIRD et al., 1974 citados por TEIXEIRA, 1995). Igualmente, a polpa de beterraba pode substituir 30% de cereais não afetando, em termos de ganho de peso, o desempenho de suínos de 60 à 120kg de peso vivo (BEORLEGUI, 1990).
Os suínos são mais hábeis em digerir hemicelulose do que celulose (STANOGIAS e PEARCE, 1985), e ainda melhor a hemicelulose de leguminosas que a das gramíneas (KEYS et al., 1970). As hemiceluloses estão presentes em média de 10 a 25% na matéria seca das forragens em muitos subprodutos industriais como as polpas de citrus e de beterraba, e apenas em 2 a 12% nos cereais e raízes (GIRGER e POCHET, 1987 citados por TEIXEIRA, 1995). Segundo TEIXEIRA (1995), a maior digestibilidade da hemicelulose deve-se a uma "sensibilidade" de algumas de suas ligações químicas à acidez gástrica, de tal forma que os produtos desta hidrólise ficam expostos à digestão intestinal.
No entanto, em talo de lucerna, a digestibilidade aparente da hemicelulose parece ser menor ao da celulose e isso ocorre devido a forte ligação físicoquímica entre a hemicelulose e lignina (STANOGIAS e PEARCE, 1985).
Um dos principais responsáveis pela grande variação da digestibilidade da fibra é a lignina. Está, não é, aparentemente, degradada pelos suínos, diminuindo a digestibilidade dos polissacarídeos não amiláceos, principalmente devido às ligações covalentes mantidas com a celulose e hemicelulose, além de não ser digerida por microrganismos presentes no intestino dos suínos (JOHNSTON et al., 2003).
STANOGIAS e PEARCE (1985) notaram uma queda da digestibilidade aparente de todos os componentes dietéticos com o aumento do consumo de dieta contendo casca de ervilha. Tal queda deve-se à uma substância anti-nutritiva – o tanino. Compostos de tanino podem afetar a digestibilidade de várias maneiras: formação de complexos estáveis de taninoproteína e tanino-celulose, inibição de enzima, inibição da flora microbiana e por reação com proteínas endógenas da mucosa intestinal.
O beta-glucano é um dos principais componentes do endosperma da cevada. O alto nível de betaglucano, mostrou reduzir o valor nutricional aumentando a viscosidade do fluído intestinal (THACKER et al.,1989) e dificultando o ataque das enzimas endógenas (JOHNSTON et al., 2003).
Taxa de passagem, tamanho da partícula e categoria animal
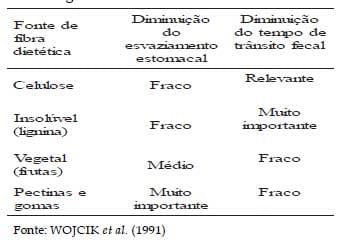
A fonte de fibra afeta a taxa de passagem e este efeito é influenciado pela capacidade dos diferentes componentes da fibra absorverem e reterem água (Quadro 1). Já para STANOGIAS e PEARCE (1985), a taxa de passagem pode estar mais relacionada com outras características físicas do alimento, como tamanho da partícula, além da quantidade ingerida de fibra e do peso do animal.
Quadro 1. Efeito da fibra dietética sobre taxa de passagem
Fazendo uma classificação arbitrária do tamanho das partículas, teríamos a seguinte distribuição: tamanho pequeno incluirá casca de aveia e talo de lucerna; tamanho médio incluirá casca de soja, milho, espiga de milho e casca de ervilha e a categoria de grande partículas compreenderia de casca de lupin e farelo de trigo (STANOGIAS e PEARCE, 1985).
FERNANDEZ e JORGESEN (1986), relataram que partículas mais finas trazem maiores benefícios para animais jovens, sendo essa influência mais acentuada com o aumento da fibra na dieta. A digestibilidade da matéria seca, energia e dos constituintes celulares do feno de alfafa aumentou , quando partículas de 6,25mm foram comparadas com partículas de 12,5mm, em dietas de fêmeas gestantes (NUZBACK et al., 1984 citados por TEIXEIRA, 1995).
Segundo VAREL et al. (1991) a idade de suínos não afetou a digestibilidade "in vitro" da fibra de alfafa. JEY e DE BARTHE (1974), relataram que alfafa pode ser adicionada de 20 à 40% para suínos na fase de crescimento e terminação, sem afetar significativamente o ganho de peso. Tal fato foi observado, também, por WOJCIK et al. (1991) com suínos de 20 à 100kg da raça Large-White que receberam até 11,85% de alfafa e garantiram um ganho de peso de 601 à 613g dia-1. Porém, CLOSE (1994), salientou que as porcas são mais aptas na utilização de fibra do que suínos desmamados ou em fase de crescimento/ terminação. A idade e peso afetam a digestibilidade da fibra, a qual aumenta com o aumento da idade, sendo a diferença entre animais jovens e adultos maior quando a fibra é pouco digerível. Animais mais velhos digerem melhor a fibra, provavelmente pelo maior desenvolvimento do ceco, maior tempo de retenção da digesta (FERREIRA, 1990 citado por TEIXEIRA, 1995) e por conter maior quantidade de bactérias fermentativas no trato (YEN et al, 2004). Fato este já constatado por VAREL e YEN (1997), ao observarem um número de bactérias celulíticas 6,7 vezes maior em suínos adultos que em suínos em crescimento.
FERNANDEZ e JORGESEN (1986) observaram que suínos de 20 à 90kg de peso vivo recebendo níveis variados de fibra bruta na dieta (3 à 16%) apresentavam menor digestibilidade à medida que os níveis de fibra aumentavam, porém um aumento da digestibilidade em todos os níveis de fibra bruta foi notado com o aumento do peso vivo dos animais.
A inclusão de alimentos fibrosos em dietas de porcas não necessariamente deprime a digestibilidade dos nutrientes. Recentemente, RENTERIA FLORES (2003) alimentou porcas gestantes com dieta a base de milho e soja (controle); milho+soja e 34% de farelo de aveia; milho+soja e 12% de palha de trigo ou milho+soja e 16% de polpa de beterraba. A digestibilidade da energia e do nitrogênio foram similares entre as dietas com farelo de aveia e controle. A digestibilidade aparente da energia e nitrogênio foi deprimida pela adição de palha de trigo, sendo a digestibilidade da dieta com polpa de beterraba intermediária. Os autores concluíram que o grau de digestibilidade depende da característica da fibra utilizada.
Alguns estudos têm evidenciado melhora no desempenho reprodutivo de porcas alimentadas com dietas ricas em fibra alimentar. GRIESHOP et al. (2001) revisaram 20 estudos citados na literatura sobre a utilização de dietas com alta fibra sobre as características reprodutivas de porcas. Treze dos 19 estudos que reportaram dados sobre o tamanho de leitegada mostraram uma melhora deste índice em fêmeas com acesso a fibra durante a gestação. Seis estudos não apresentaram diferenças ou uma piora no referido índice zootécnico. A magnitude das respostas positivas foram representadas em um intervalo de 0,1 a 2,3 leitões nascidos vivos/leitegada. A fibra durante a gestação melhorou a longevidade das matrizes em 4 dos 8 estudos que consideraram esta característica.
Este efeito positivo em relação ao tamanho da leitegada pode estar relacionado a um menor aporte de energia líquida, proporcionado pela fibra, logo após a cobertura o que diminuiria as perdas embrionárias (JOHNSTON et al., 2003). Além disso, o uso de fibra também pode apresentar um reflexo positivo na diminuição de comportamentos estereotipados em fêmeas confinadas, diminuindo o estresse por aumentar seu nível de saciedade JOHNSTON et al., 2003).
Degradação da fibra: sítios
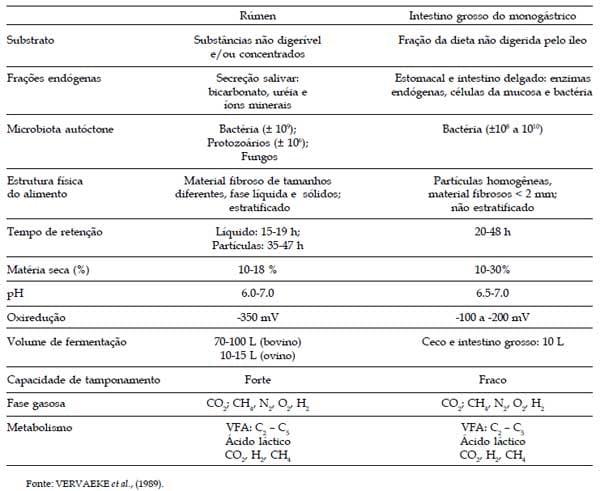
Há um reconhecido conceito de que o mecanismo da digestão da fibra no ceco e intestino grosso de monogástrico, especialmente suínos, é muito semelhante à fermentação que ocorre no rúmen (VERVAEKE et al., 1989). Admite-se semelhanças nas características da fermentação quanto ao pH e composição de ácidos graxos voláteis (VERVAEKE, 1978). Porém, é claro, que há muitas diferenças entre a fermentação do rúmen e do intestino, diferenças estas que podem ser importantes para uma específica aproximação e interpretação da fibra em ambos os sistemas. Tais diferenças e similaridades são sumarizadas no Quadro 2.
ZEBROWSKA (1991) observou que a digestibilidade da fibra bruta e da fibra em detergente neutro no intestino delgado foi de 42%; 80% para centeio e 34%; 72% para trigo, sendo maior do que a de cevada (15%; 62%) e farelo de trigo (23%; 45%).
Estudos com suínos canulados revelaram existir alguma degradação da fibra anteriormente ao ceco, especialmente com algumas leguminosas facilmente fermentáveis e polpa de citrus e de beterraba (DIERIK et al., 1989).
KASS et al. (1980) reportaram digestão da ordem de 38,2% da hemicelulose contida na alfafa, anteriormente ao íleo, em suínos e MILLARD e CHESSON (1984) confirmaram essa possibilidade pela identificação de bactérias anaeróbias celulolíticas e pectinolíticas na digesta presente no íleo de suínos.
Quadro 2. Semelhanças e diferenças entre as condições do rúmen e do intestino grosso do monogástrico
Por outro lado, KEYS e DE BARTHE (1974) citados por TEIXEIRA (1995), constataram que a maior parte celulose e da hemicelulose do capim bermuda (Cinodon dactylon-L) parecia ser digerido no intestino delgado desses animais.
Degradação da fibra: microorganismos participantes
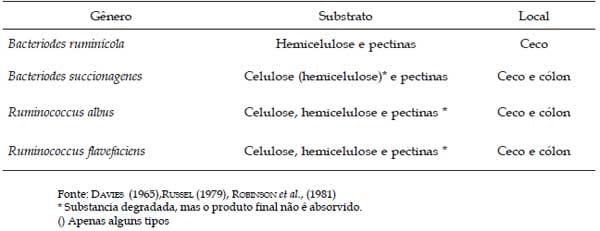
No Quadro 3 são apresentados os microorganismos participantes da degradação da fibra, bem como o substrato de ação e localização.
A parede celular dos polissacarídeos são degradados em maior ou menor extensão no ceco e cólon por uma diversificada população microbiana, para a qual eles são a maior fonte de carbono. As condições de biotope (baixo tempo de trânsito, pH, umidade e temperatura) favorecem essa degradação (FONTY e GOUET, 1989).
JENSEN e JORGENSEN (1996), relataram que a maior atividade microbiana foi encontrada no ceco e no cólon proximal, e que a atividade microbiana no estômago e intestino foi maior em suínos que receberam dieta com alto teor de fibra do que o de baixo teor de fibra.
VAREL et al. (1984), observaram que a amostra fecal coletada de porcas cruzadas alimentadas com dieta rica em fibra (35% de farinha de alfafa desidratada) apresentou um número de bactéria celulolítica maior do que amostras de suínos alimentados com dieta à base de milho e soja.
Quadro 3. Principais bactérias que degradam a parede celular vegetal presente no trato digestivo de suínos.
VAREL e POND (1985), analisando amostra fecal de porcas gestantes alimentadas com vários níveis de fontes de fibra (controle grão e soja; 20% de espiga de milho e 40% e 96% de farinha de alfafa), verificaram que o total de bactérias celulolíticas, também, aumentaram com o aumento da concentração de fibra ou parede celular da planta na dieta. Porém, isso depende da fonte fibrosa, pois quando farinha de alfafa foi oferecida houve um aumento significativamente maior no número total de bactérias celulolíticas quando comparado com a inclusão de 20% de espiga de milho.
Segundo STAGIONAS e PEARCE (1985) a possível queda da digestibilidade da celulose da casca de milho, espiga de milho e casca de aveia se deve ao baixo teor de nitrogênio nessas fibras. Salientaram, ainda, que é razoável assumir que a fermentação pela bactéria no ceco e no intestino grosso de suínos, como no rúmen, requer adequada concentração de nitrogênio para a ótima atuação.
O número de bactérias, celulolíticas e hemicelulolíticas, aumentaram dentro de 3 dias após o início do fornecimento de dieta de alta fibra (FONTY e GOUET, 1989). VAREL et al. (1991), relataram que a população bacteriana parece adaptar-se à dieta com alta fibra dentro de 14 dias.
A adaptação da bactéria celulolítica ocorre mais rapidamente no genótipo magro do que no obeso e o número em suínos magros alimentados com dieta de alta fibra aumenta 17 vezes, enquanto aumento não significativo foi observado em suínos magros alimentados com dieta de baixa fibra (FONTY e GOUET, 1985).
A microbiota intestinal dos suínos contém alta atividade de bactérias de espécies celulolíticas e hemicelulolíticas, a qual incluem Fibrobacter succinogenes, Ruminococcus albus, Ruminococcus flavefaciens, Butyrivibrio spp. e Prevotella ruminicola, além do Clostridium herbivorans uma bactéria de alta atividade celulolítica presente no intestino grosso (VAREL e YEN, 1997).
Ácidos graxos voláteis produzidos
A quebra microbiana da parede celular de polissacarídeos e subsequente fermentação resulta na produção de uma mistura de ácidos graxos voláteis, compostos, principalmente de ácido acético, propiônico e butírico (STAGIONAS e PEARCE, 1985).
Segundo TEIXEIRA (1995), a concentração de ácidos graxos voláteis presente no trato gastrointestinal está relacionado com o tempo após a ingestão, e a composição da dieta. A produção de ácidos graxos voláteis no intestino grosso dos suínos é influenciado, principalmente pelo tipo e nível de fibra da dieta. STAGIONAS e PEARCE (1985), observaram que a concentração total de ácidos graxos no intestino grosso aumentou linearmente com o aumento de fibra em detergente neutro ingerida, relataram ainda que os efeitos do nível e da fonte da fibra da dieta na proporção molar dos ácidos graxos da digesta foi variável.
O suprimento energético oferecido pelos ácidos graxos voláteis é, ainda, muito discutido.POND (1989), sugeriu que 5 à 25% da energia de mantença requerida para o crescimento de suíno é fornecido pelos ácidos graxos voláteis; assim como CLOSE (1994), o qual relata que a fibra pode contribuir em mais de 30% da energia requerida. Já DIERICK (1990), diz que os ácidos graxos voláteis irão fazer apenas uma pequena, mas significante contribuição na energia total utilizável para suínos em crescimento/terminação.
O aumento dos níveis de fibra da dieta resultam num significativo aumento da razão acetato/ propionato. A adição de 27,3% e 52% de feno de alfafa em dietas para suínos, resultou em significativo aumento de porcentagens de ácido acético em relação ao propiônico. Também, 35% de feno de alfafa provocou uma diminuição de propionato produzido, aumentando a relação acetato-propionato (TEIXEIRA, 1995). ZEBROWSKA (1991) relatou que a produção de ácidos graxos voláteis na digesta do ceco foi maior quando suínos foram alimentados com centeio do que com trigo, cevada ou farelo de trigo.
STAGIONAS e PEARCE (1985), relataram que ácidos graxos voláteis são facilmente absorvidos pela mucosa intestinal e que as maiores concentrações de ácidos graxos voláteis no conteúdo do ceco e da porção proximal do intestino grosso foram alcançados 2-4 horas após a alimentação.
O aumento da concentração de ácidos graxos voláteis aumenta com o consumo de fibra em detergente neutro pelos suínos, sugerindo que maiores quantidades de substrato fermentativo são colocados disponíveis para os microorganismos. Se tal aumento de quantidade de ácidos graxos voláteis são absorvidos pelo intestino e metabolizados em energia, então, presumivelmente, mais energia poderia ficar disponível para suínos com o aumento de consumo de fibra detergente neutro na dieta (STAGIONAS e PEARCE, 1985).
Na utilização de fibra altamente fermentável (20% de polpa de beterraba) o suprimento de energia pelos ácidos graxos voláteis foi 28% como parte da energia de mantença (VERVAEKE et al., 1989).
No entanto, é importante ressaltar que a eficácia de transformação de ácidos graxos voláteis em energia é inferior à eficácia de aproveitamento de carboidratos solúveis, como a glicose (TEIXEIRA, 1995). LOW(1985) demonstrou, em suínos em crescimento, que a eficácia de utilização dos ácidos graxos voláteis corresponde à 75% da eficácia de transformação de glicose em energia.
Infuência da fibra: digestibilidade e absorção dos nutrientes
A digestibilidade aparente da matéria seca, nitrogênio, energia e de outros componentes não fibrosos, é inversamente relacionada com a proporção de fibra na dieta, ou a quantidade de consumo de fibra (STANOGIAS e PEARCE, 1985). Tal afirmação é alicerçada por LACHANGE (1992), o qual relata que aumentado o teor de fibra da dieta, diminui significantemente a digestibilidade aparente da proteína bruta, energia, lipídeos, fibra bruta, fibra detergente neutro, fibra detergente ácida e hemicelulose e por HARTOG et al.(1988), que observaram uma queda na digestibilidade ileal e fecal da matéria orgânica com a inclusão de celulose.
Muitos estudos tem sido realizados utilizandose suínos para determinar o efeito do nível e fonte de fibra sobre a digestibilidade ileal da matéria seca e energia. Tal fato é resultado direto da substituição de fonte de carbono altamente digerível por uma de baixa digestibilidade (SAUER et al., 1991).
Teoricamente, a fibra dietética pode reduzir a digestibilidade da proteína e de aminoácidos por meio de estímulo da produção de proteína de origem bacteriana, através da adsorção de aminoácidos e peptídeos para a matriz da fibra e pelo aumento de secreção da proteína endógena. Além disso, temse constatado que a inclusão de fibra na dieta resulta em aumento da descamação da mucosa intestinal e incremento da produção de muco, levando ao aumento da perda de aminoácidos endógenos (POZZA et al., 2003).
SAUER et al..(1991), relataram uma significante diminuição da digestibilidade ileal dos aminoácidos quando 5% ou 10% de celulose foi incluído na dieta. Tal observação, também, foi verificada por HARTOG et al. (1988), que relataram uma queda da digestibilidade dos aminoácidos (isoleucina, lisina, fenilalanina, treonina, valina, alanina, ácido aspártico, ácido glutâmico e tirosina) com adição de 5% de palha.
A inclusão de 6% de metil-celulose diminuiu a digestibilidade ileal aparente de nitrogênio de 76 para 48%. A inclusão de 7,5% de pectina em uma dieta formulada à base de farelo de soja diminuiu a digestibilidade média ileal de aminoácidos essenciais de 83,9% para 70,6% (SAUER et al.., 1991).
POZZA et al. (2003) verificaram que o aumento do níveis de fibra dietética, provenientes da inclusão de casca de arroz à dieta isenta de proteína, resultou em aumento da perda endógena dos aminoácidos essenciais, com exceção da glicina.
A utilização de fibra dietética na alimentação de suínos também pode afetar a absorção de minerais.
Pectina foi introduzida à um nível de 5% ocasionando uma queda na digestibilidade ileal de sódio, potássio, magnésio, cálcio e potássio (HARTOG et al., 1988).
MOORE et al.. (1986), relataram que 10% de casca de aveia diminui o balanço de zinco e cálcio de suínos alimentados durante 7 dias, porém este não mostrou efeito sobre o balanço mineral em suínos alimentados com a dieta por 6 ou 26 dias.
NEWTON et al.(1983), mostraram que 20% de farelo de trigo aumenta absorção de ferro, mas reduz absorção de zinco em suínos alimentados por um período de 80 dias, porém não afeta a absorção aparente de minerais nos mesmos suínos quando recebem dieta por um período de 14 dias.
A absorção aparente de sódio foi reduzido quando suínos receberam dieta contendo 20% de farinha de alfafa, porém a absorção foi maior do que para os suínos alimentados com dieta contendo farelo de soja. Tal fato ocorreu, primeiramente, devido ao alto consumo de sódio. Farelo de soja, 15% de casca de aveia, 15% de casca de soja e 20% de farinha de alfafa, aumentaram o consumo de potássio, porém apenas casca de soja e farinha de alfafa aumentaram a absorção de potássio (MOORE et al., 1988).
Dieta com 15% de casca de aveia aumentou a absorção de cobre; dieta com 15% de casca de soja e dieta com 20% de farinha de alfafa aumentaram a absorção e retenção de ferro (MOORE et al., 1988).
Há uma diversidade de resultados obtidos resultante dos diferentes teores de mineral e consumo de fontes de fibra e variação do animal, bem como, a diferença na composição da fonte fibrosa.
Influência da fibra na morfologia intestinal
POND (1989), relatou que suínos alimentados com dieta com alto teores de fibra tem intestino delgado mais pesado do que aqueles que receberam alimento com baixo teor de fibra. Tal afirmação já havia sido observada por POND et al. (1988) no qual o peso relativo (porcentagem do peso vivo) do fígado, coração, estômago cheio e intestino delgado, bem como, o ceco e cólon foram maiores em suínos alimentados com dieta com alto teor de fibra do que os que receberam dieta com baixo teor de fibra. Essa observação tem um significado especial quando considera- se a alta correlação existente entre o peso do órgão cheio e a produção de calor (POND, 1989).
O proporcional aumento do tamanho e peso do trato gastrointestinal pode influenciar diretamente nas exigências de manutenção relacionadas a energia e aminoácidos, pois existe maior demanda energética advinda da alta taxa de renovação celular presente neste órgãos (JOHNSTON et al., 2003).
MEULEN et al. (1991) observaram que a quantidade de digesta do estômago e do intestino delgado aumentou pela inclusão de fibra, particularmente, de farelo de trigo GOMES et al. (1998b) observaram alterações morfológicas dos órgãos digestivos de leitões em fase de recria quando estes foram alimentados com dieta contendo 10% de feno de Tifton (Cynodon dactilon).
Evidências de danificação do epitélio foi observado em alguns suínos alimentados com dietas com alto teor de fibra, sugerindo que alguns animais são susceptíveis á mudança morfológica do intestino quando recebem dieta com alto teor de fibra (MOORE et al., 1988).
JIN et al. (1994) demonstrou que dietas com conteúdo elevado de fibra insolúvel proveniente da palha do trigo aumentou a proliferação celular das criptas do jejuno e cólon e aumentou a taxa de morte de células epiteliais do jejuno e íleo de suínos em crescimento. A proliferação de células epiteliais é suportada pelo butirato o qual é produzido na fermentação de alimentos fibrosos (MONTAGNE et al.,2003).
Efeitos semelhantes foram observados por Gomes et al. (1998a) quando o incremento de baixo nível de FDN (8%) na ração de leitões em recria promoveu alterações e atuou sobre o epitélio dos mesmos, de maneira a elevar a área celular, bem como elevar a produção e liberação de muco pela mucosa intestinal. Em suínos em fase de terminação consumindo ração com 8% FDN foi observado aumento no percentual da área ocupada pelas glândulas do epitélio do ceco e redução no percentual da área ocupada pelas células caliciformes (GOMES et al., 2001).
Em vista destes resultados podemos concluir que o oferecimento de dietas compostas por ingredientes fibrosos para suínos em suas diversas categorias proporciona uma provável adaptação histo-fisiológica e mecânica às ações abrasiva e fermentativa exercida pela fração fibra dietética (GOMES et al., 2001).
Influência do tipo animal na digestibilidade da fibra
PEKAS et al. (1983) relataram haver diferenças genéticas entre suínos na fase de crescimento e terminação e sua respostas diante dietas de alto teor de fibra. O trato gastrointestinal de suínos geneticamente magro eram mais leves do que os dos suínos tipo obeso. Porém, segundo COEY e ROBINSON (1954) e BOHMAN et al. (1955) citados por POND et al. (1988) a hipertrofia associada com o alto teor de fibra da dieta é maior em suínos do tipo magro.
VAREL et al. (1982) sugeriram uma diferença genética entre suínos do tipo obeso e magro com relação a sua microbiota celulolítica e, com isso, a sua habilidade de adaptação à dietas com alto teor de fibra.
VAREL et al. (1988) ofereceram dieta contendo 80% de alfafa e verificaram que o número de bactérias celulolítica para o tipo obeso, magro e contemporâneo foi maior (23x10 ; 51,6x10 e 37,2x10, respectivamente) e que o tempo de passagem da digesta foi maior para o tipo obeso.
POND et al. (1988) observaram que a restrição de energia reduz a gordura do toucinho nos suínos tipo obeso, magro e contemporâneo, porém, o tipo obeso, em relação aos outros genótipos, apresenta maior toucinho, menor músculo longissimus dorsi, coração, estômago e cólon.
Mais recentemente YEN et al. (2004) estudaram o potencial de utilização de alimento fibroso das raças cruzadas criadas no ocidente (Duroc x Branco) e do suíno Meishan de origem chinesa, os autores verificaram não haver diferença na habilidade de digerir fibra e na produção de AGV entre os dois genótipos.
Influência da temperatura ambiente na digestibilidade da fibra
NIENABER e HAHN (1983), observaram que suínos alimentados "ad libitum" tendem a consumir exigências energéticas para mantença e crescimento quando mantidos em ambiente termoneutro ou moderadamente frio (5 à 8°C abaixo LCT), mas não quando em ambiente quente.
Devido ao fato da fibra ter uma alta energia, a qual perde resultando num incremento calórico, o valor nutricional desta pode ser afetado pela temperatura ambiente. O calor gerado com a digestão do nutriente e do metabolismo tem uma função utilizável em ambiente frio através da minimização da oxidação de outros nutrientes e tecidos para a produção de calor. A adição de 10% de farinha desidratada de alfafa diminui em apenas 1% o ganho de peso diário em ambiente frio (8°C) quando comparado com 3% e 5% em ambientes com 22,5 e 35°C respectivamente. A eficiência alimentar diminui em 1%, 7% e 10% em ambiente frio, médio e quente, respectivamente (ATAHLY e CROMWEL, 1986).
HATA e KOTZUMI (1996) comparando suínos recebendo dieta com 2,5% (baixa ) e 7,7% (alta) de fibra alojados em ambientes de alta temperatura (20°C) e baixa (5°C) concluíram que a digestibilidade da dieta de alta fibra foi de 5% à 20°C enquanto que à 5°C houve uma queda na digestibilidade da matéria seca, proteína bruta, energia dos nutrientes.
Por outro lado, JORGENSEN et al. (1996), relataram que suínos alojados em ambiente de baixa temperatura (13°C) tem menor ceco do que os alojados em ambiente com alta temperatura (23°C) e não encontraram efeito significativo na inter-relação fibra x temperatura ambiente sobre a energia metabólica, apesar da queda de 13°C para 23°C causar uma queda no incremento calórico de 2,9MJ/suíno dia.
FASSHAUER (1993), relatou que a inclusão de aditivo antimicrobiano, ácido cítrico na quantidade de 10g kg-1 de ração aumenta a digestibilidade da fibra bruta.
A adição de bentonato de sódio e beta-glucanase em dietas com cevada aumentou o coeficiente de digestibilidade da energia em comparação aos animais que receberam dieta adicionando somente beta-glucanase ou nenhum dos dois aditivos (THACKER et al., 1989).
CONSIDERAÇÕES GERAIS
A utilização de alimentos alternativos, ricos em fibra, para suínos deve ser avaliada com especial atenção, pois sua produção é garantida, uma vez que a maioria desses alimentos são resíduos da própria agricultura direcionada ao homem. Além disso, o seu baixo custo permite a inclusão na produção de suínos, com barateamento das rações.
A capacidade do suíno degradar alimentos fibrosos dependerá de alguns fatores, os quais devem ser considerados quando se pretende introduzir fontes fibrosas na alimentação, dentre estes destacamos:
Quanto ao tipo de fibra: os alimentos ricos em celulose e lignina são menos digeridos pelos suínos, enquanto que os ricos em hemicelulose são mais digeríveis. Além da composição da fibra, fatores antinutricionais, como o tanino presente na casca de ervilha e o beta-glucano um dos componentes principais do endosperma da cevada, mostraram reduzir a digestibilidade. Alimentos ricos em pectinas como a polpa de beterraba, polpa de citrus, casca da soja e sementes de leguminosas são bem digeridas pelos suínos.
Quanto a taxa de passagem: esta varia de acordo com a composição, tamanho de partícula, quantidade de ingestão de fibra e do peso do animal.
Quanto ao tamanho da partícula: partículas menores trazem mais benefícios para animais jovens e adultos quanto a digestibilidade.
Quanto à idade: as opiniões são contraditórias, porém predomina que os animais adultos e porcas são mais hábeis para digerir fibra.
A digestão da fibra pelos suínos deve-se à flora microbiana do ceco e cólon desses animais, a qual requer adequada concentração de nitrogênio para a sua ótima atuação.
Alterações da morfologia intestinal de suínos alimentados com fibra foi observado com um aumento dos órgãos viscerais.
O produto final da fermentação da fibra são os ácidos graxos voláteis, os quais suprem em média 30% da energia total requerida.
A fibra influência negativamente a digestibilidade de outros nutrientes, como proteína, energia, e até mesmo dos próprios componentes da fibra e na absorção dos minerais.
Animais alimentados com fontes fibrosas apresentam carcaças mais magras e de melhor qualidade. Animais geneticamente do tipo magro adaptamse mais rapidamente à dieta fibrosa do que os do tipo obeso. A resposta a dietas com fibra é influenciada pela temperatura ambiente.
A utilização de alimentos fibrosos para porcas visa, também, controlar possíveis problemas de excessiva ingestão alimentar e o estresse advindo do confinamento.
REFERÊNCIAS BIBLIOGRÁFICAS
BEORLEGUI, C. B. Utilization de las pupas de remolacha y citrus en piensos de ruminantes y monogastricos. Madrid: Universidad Politechica de Madrid/ETSIA, 1990. 36 p.
BRIOLLOUET, J. M.; CARRE, B. Composition of cells walls from cotyledons of Pisum sativum, Vicio jaca and Glicine max. Phytochemistry, Oxford, v.22, p.841-847, 1983.
CLOSE, W. H. Fibrous diets for pigs. Pig News Information,Oxon, v. 15, p. 65, 1994.
DAVIES, M. E. Cellulolytic bacteria in some ruminants and herbivores as shown by fluorescent antibody. Journal of Genetic Microbiology,London, v. 39, p. 139-141, 1965.
DIERICK, N. A. et al. Approach to the energetic importance of fiber digestion in pigs. 1. Importance of fermentation in the overall energy suplly. Animal Feed Science and Technology, Amsterdam, v.23, n. 1-3, p.141- 167, 1989.
FAO. New Feed Resoucers. Rome: FAO Production and Health, 1977.
FASSHAUER, V. Effect of antimicrobial feed additives on apparent precaecal digestibility of nitrogen and minerals in the pig. Hannover: Institut fur Tierernahrung, Tierarztliche Hochschule Hannover,1993. 88 p.
FERNANDEZ, A.G.; JORGESEN, N.H. Comparative digestibility experiments with growing pigs and adult sows. Animal Production, Edinburgh, v.43, n.1, p.127- 132, 1986.
FONTY, G.; GOUET, P.H. Fiber degranding microorganisms in the monogastric digestive tract. Animal Feed Science and Technology, Amsterdam, v.23, p.91-107, 1989.
GOMES, J.D.F. et al. Efeitos do incremento de fibra em detergente neutro (FDN) sobre a histologia intestinal de suínos em fase final de terminação. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38., 2001, Piracicaba. Anais... Piracicaba: Sociedade Brasileira de Zootecnia, 2001. CD-ROM.
GOMES, J.D.F. et al. Efeitos do incremento de fibra em detergente neutro sobre a histologia do epitélio intestinal de suínos em recria. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35, 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, 1998a .CD-ROM.
GOMES, J.D.F. et al. Efeitos do incremento de fibra em detergente neutro sobre o desempenho; características de carcaça e morfologia do trato gastrointestinal de suínos em recria. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35, 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, 1998b. CDROM.
GRIESHOP, C.M.; REESE, D.E.; FAHEY Jr, G.C. Nonstarch polysaccharides and oligosaccharides in swine nutrition. In: LEWIS, A.J, SOUTHERN, L.L. (Ed). Swine Nutrition. Boca Raton: Ed. CRC Press, 2001. p.107
HARTLEY, R. D.; JONES, E. C. Phenolic components and degradability of cell walls of grass and legume species. Phytochemistry, Oxford, v. 16, p. 1531-1534, 1977.
HARTOG DE, L. A. et al. The effect of including various structural polyssaccharides in pig diets on ileal and faecal digestibility of aminoacids and minerals and minerals. Livestock Production Science, Amsterdam, v.18, p.157- 170, 1988.
HATA, H.; KOIZUMI, T. Effect of cold on digestibility in pigs in relation to dietary fiber content and feeding level. Pig News Information,Oxon, v. 17, n.1, p.79, 1996.
JENSEN, B. B.; JORGENSEN, H. Effect of dietary fiber on microbial activity and microbial gas production in various regions of the gastrointestinal tract of pigs. Pig News Information,Oxon, v.17, n.13, p.308, 1996.
JIN, L. et al. Effects of dietary fiber on intestinal growth, cell proliferation, and morphology in growing pigs. Journal of Animal Science, Champaign, v.72, p.2270- 2278, 1994.
JOHNSTON, L.J. et al. Feeding by-products high in concentration of fiber to non-ruminants. In: THIRD NATIONAL SYMPOSIUM ON ALTERNATIVE FEEDS FOR LIVESTOCK AND POULTRY HELD IN KANSAS CITY. Proceedings... Kansas City: 2003. p. 1-26.
JORGENSEN, N. H.; ZHAO, X.; EGGUM, B.O. The influence of dietary fiber and environmental temperature on the development of the gastrointestinal tract, digestibility, degree of fermentation in the hind-gut and energy metabolism in pig. Pig News Information, Oxon, v.17, n. 2, p. 197, 1996.
KEYS Jr, J. E.; VAN SOEST, P.J.; YOUNG, E.P. Effect of increasing dietary cell wall content on the digestibility of hemicellulose and cellulose in swine and rats. Journal of Animal Science, Champaign, v. 31, n. 6, p. 1172-1177, 1970.
LACHANCE, B.; FEURIER, C.; VINET, C. In the growing pig nutrient digestibility may be influenced by the level of dietary fiber and the amount of whey. Bulletin Agriculture Canada, Research Branch, n.12, p.20-23, 1989.
MONTAGNE, L.; PLUSKE, J.R.; HAMPSON, D.J. A review of interactions between fiber and the intestinal mucosa, and their consequences on digestive healthin young non-ruminant animals. Animal Feed Science and Technology, Amsterdam, v.108, p.95-117, 2003.
MOORE, R. J.; KORNEGAY, E. T. Fiber digestibility and mineral utilization in growing pig as influenced by fiber source, mineral level and duration of feeding. Nutrition Report International, Los Altos, v. 36, n. 6, p. 1237-1240, 1987.
MOORE, R. J. et al. Growth nutrient utilization and intestinal morphology of pigs fed high-fiber diets. Journal of Animal Science, Champaign,v.66, p. 1570-1579, 1988.
MOORE, R. J.; KORNEGAY, E. T.; LINDEMANN, M. D. Effect of dietary oat hulls or wheat bran on mineral utilization in growing pigs fed diets with or without salinomycin. Canadian Journal Animal Science,Ottawa, p. 256-267, 1986.
NATIONAL RESEARCH COUNCIL. Nutrients requirements of Swine. 10.ed. Washington National Academic Science, 1998. 189 p.
NIENABER, J. A.; HAHN, G. L. Performance of growingfinishing swine in response to the thermal environment. St. Joseph: American Society Agricultural Enginnering , 1983. ( Paper MRC 83-137)
NOBLET, J.; PEREZ; S. Prediction of digestibility of nutrients and energy values of pig diets from chemical analysis. Journal of Animal Science, Champaign, v. 71, p. 3389-3398, 1993.
PEKAS, J. C.; YEN J. T.; POND, W. G. Gastrointestinal, carcass and performance traits of obese versus lean genotype swine: Effect of dietary fiber. Nutrition Reports International, Los Altos, v. 27, p. 259, 1983.
POND, W. G.; JUNG, H. G.; VAREL, V. H. Effect of dietary fiber on young adult genetically lean, obese and contemporary pigs: body weight, carcass measurements, organ weight and digesta content. Journal of Animal Science, Champaign, v.66, p. 699-706, 1988.
POND, W. G. Plant fiber utilization by pigs. Pig News Information,Oxon, v.10, n.1, p. 13-15, 1989.
POZZA, P.C. et al. Avaliação da perda endógena de aminoácidos, em função de diferentes níveis de fibra para suínos. Revista Brasileira de Zootecnia, Viçosa, v.32, n.6, p. 1354-1361, 2003.
RENTERIA FLORES, J.A. Effects of soluble and insoluble dietary fiber on diet digestibility and sow performance. 2003. (PhD. Dissertation)- University of Minnesota, St. Paul, 2003.
ROBINSON, I. M.; ALLISON, M. J.; BUCKLIN, J. A. Characterization of the cecal bacteria of normal pigs. Applied Environmental Microbiology, Washington, v. 41, p. 950-955, 1981.
RUSSELL, E. G. Types and distribution of anaerobic bacteria in the large intestine of pigs. Applied Environmental Microbiology, Washington, v. 37, p.187- 193. 1979.
SAUER, W. C.; OZIMEC, L. Digestibility of amino acids in swine: results and their practical applications. Livestock Production Science, Amsterdam, v.15, p.367-388, 1986.
SAUER, W. C. et al. The effect of source of fiber on ileal and fecal amino acid digestibility and bacterial nitrogen excretion in growing pigs. Journal of Animal Science,Champaign, v.69, n. 10, p. 4070-4077, 1991.
SCHOENHERR, W. D.; STAHLY, T. S.; GROMWELL, G. L. The effects of dietary fat or fiber addition on energy and nitrogen digestibility in lactating, primiparous sows housed in a warm or hot environment. Journal of Animal Science, Champaign,v.67, p.473-481, 1989.
SILVA, D. J. Análise de Alimentos (métodos químicos e biológicos). 2. ed. Viçosa: UFV, 1990. 165 p.
SMITH, G. S.; NELSON, A. B.; BOGGINO, E. J. A. Digestibility of forrages in vitro as affected by content of silica. Journal of Animal Science, Champaign, v. 33, p. 466-471, 1971.
STAHLY, T. S.; CROMWELL, G. L. Responses to dietary addition of fiber (alfafa meal) in growing pigs housed in a cold, warm or hot thermal environment. Journal of Animal Science, Champaign, v. 63, p.1870-1876, 1986.
STAGONIAS, G.;PEARCE, G. R The digestion of fiber by pigs. 1. The effects of amount and type of fiber on apparent digestibility, nitrogen balance and rate of passage. British Journal Nutrition, London, v.53, p.513-530, 1985.
TEIXEIRA, E. W. Utilização de alimentos fibrosos pelos suínos. Zootecnia, Nova Odessa, v. 33, n. 1, p. 19-27, 1995.
THACKER, P. A.; CAMPBELL, G. L.; GROOTWASSINK, J. W. D. The effect of sodium bentonite on the performance of pigs fed barley-based diets supplemented with betaglucanase. Nutrition Reports International, Los Altos, v. 40, n.3, p. 613-619, 1989.
THEANDER, O.; UDÉN, P.; AMAN, P. Acetyl and phenolic acid substituents in timo by of different maturity and after digestion with rumen microorganisms or a commercial cellulase. Agriculture and Environment ,Amsterdam, v. 6, p. 127-133, 1981.
THEANDER, O. et al. Plant cell wall and monogastric diets. Animal Feed Science and Technology, Amsterdam, v. 23, p. 2,5-25, 1989.
TUNGLAND, B.C.; MEYER, D. Nondigestible oligo and polysaccharides (dietary fiber): their physiology and role in human and health food. Comprehensive Reviews in Food Science and Food Safety, v.1, p.73-77, 2002.
VAN SOEST, P. J. Chemical properties of fiber in concentrate feeds stuffs. Proceedings of the Cornell Nutrition Conference, p. 17-21, 1969.
VAN SOEST, P. J. The role of silicon in the nutrition of plants and animals. Proceedings of the Cornell Nutrition Conference, p. 103-109, 1970.
VAN SOEST, P.J. ; JONES, L. H. P. Effect of silica in forages upon digestibility. Journal of Dairy Science, Champaign, v. 51, p. 1644-1648, 1968.
VAREL, V. H.; FRYDA, S. J.; ROBINSON, I. M. Cellulolytic bacteria from pig large intestine. Applied and Environmental Microbiology, Washington, v. 47, p. 219- 220, 1984.
VAREL, V. H.; JUNG, H. G.; POND, W. G. Activity of fiber degrading microorganisms in lean, obese and contemporary swine genotypes. Swine Research Program Report, n.3, p. 64-65, 1989.
VAREL, V. H.; JUNG, H. G.; POND, W. G. Effects of dietary fiber of young adult genetically lean, obese and contemporary pigs: Rate of passage, digestibility and microbiological data. Journal of Animal Science, Champaign, v. 66, p. 707-712, 1988.
VAREL, V. H. et al. Influence of high-fiber on bacterial populations in gastrointestinal tracts of obese and lean genotypes pigs. Applied and Environmental Microbiology, Washington, , v. 44, p. 107, 1982.
VAREL, V.H.; YEN, J.T. Microbial perspective on fiber utilization by swine. Journal of Animal Science,Champaign, v.75, p.2715-2722, 1997.
VERVAEKE, I. J. et al. Approach to the energetic importance of fiber digestion in pigs. II An experimental approach to hindgut digestion. Animal Feed Science and Technology, Amsterdam, v. 23, p. 169-194, 1989.
WAITE, R.; GORROD, A. R. N. The structural carbohydrats of grasses. Journal of Science and Food Agriculture, Oxford, v.10, p.308-316, 1959.
WOJCIK, S. et al. Effect of fibrous fooder addition to rations on production results of fattening pigs. Roczniki Nauk Rolniczych, Zootecchniczna, Warsaw, v. 106, n. 1- 2, p. 39-49, 1991.
YEN, J.T.; VAREL, V.H.; NIENABER, J.A. Metabolic and microbial responses in western croosbred and Meishan growing pigs fed a high-fiber diet. Animal Science,London, v.82, p.1740-1755, 2004.
ZEBROWSKA, H. Digestion of starch and fiber of cereal feeds in different section of the digestive tract of the pig. Pig News Information, Oxon, v. 12, n. 1, p. 154, 1991.
***O trabalho foi originalmente publicado no Boletim da Indústria Animal (BIA), do Instituto Zootecnia (IZ/APTA), da Secretaria de Agricultura e Abastecimento do estado de São Paulo, Brasil.