Emissões de metano na pecuária: conceitos, métodos de avaliação e estratégias de mitigação. Parte I
Publicado: 24 de abril de 2013
Por: Fernanda Samarini Machado, Luiz Gustavo Ribeiro Pereira, Fernando César Ferraz Lopes, Mariana Magalhães Campos e Mirton José Frota Morenz da Embrapa Gado de Leite, MG; Roberto Guimarães Júnior da Embrapa Cerrados, DF, e Alexandre Vieira Chaves da University of
Sydney, Sydney, Austrália.
Introdução
O crescimento da população mundial e do seu poder aquisitivo tem promovido aumento acentuado da demanda por alimentos de origem animal. Projeta-se um aumento na produção mundial de carne de 229 milhões de toneladas em 1999-2001 para 465 milhões de toneladas em 2050, e na produção de leite de 580 para 1.043 milhões de toneladas nesse mesmo período (FAO, 2006). O Brasil ocupa posição de destaque na produção pecuária, sendo importante fornecedor de proteína animal para a população mundial. Atualmente o país possui o maior rebanho comercial bovino, com 171,6 milhões de cabeças (IBGE, 2009) e detém, aproximadamente, 20% do mercado da carne (USDA, 2009), sendo o 6º maior produtor de leite (FAO, 2010).
Apesar da reconhecida importância da agropecuária na produção de alimentos e geração de renda, atualmente muito se discute sobre o impacto ambiental das atividades pecuárias e agrícolas, principalmente relativo às mudanças climáticas. A pecuária brasileira, em especial, vem sendo criticada por emitir quantidades significativas de gases de efeito estufa (GEE). Tal crítica tem sido fundamentada nos baixos índices zootécnicos verificados em sistemas de exploração animal baseados em pastagens degradadas ou que se encontram abaixo do seu potencial de produção. A ineficiência desse modelo de exploração tem gerado maiores quantidades de GEE por quilo de carne e/ou de leite produzidos (IPCC, 2007).
Dentre os vários GEE, a agropecuária contribui de forma significativa com a emissão de três deles: metano (CH4), dióxido de carbono (CO2) e óxido nitroso (NO2). O gás metano apresenta potencial de aquecimento global 25 vezes maior que o CO2 e tempo de vida na atmosfera de 9 a 15 anos, sendo sua taxa de crescimento anual de 7,0% (IPCC, 2006). A produção de metano resulta da fermentação anaeróbica da matéria orgânica em ambientes alagados, campos de arroz cultivados por irrigação de inundação, fermentação entérica, tratamento anaeróbico de resíduos animais e queima de biomassa.
O metano produzido em sistemas de produção de bovinos origina-se, principalmente, da fermentação entérica (85 a 90%), sendo o restante produzido a partir dos dejetos destes animais. Do metano produzido por fermentação entérica no rúmen, 95% é excretado por eructação, e daquele produzido no trato digestivo posterior, 89% é excretado através da respiração e apenas 11% pelo ânus (MURRAY et al., 1976). O metano derivado da fermentação entérica de ruminantes representa cerca de ¼ das emissões antropogênicas desse gás (WUEBBLES E HAYHOE, 2002). Bovinos produzem de 150 a 420 litros de CH4 por dia e ovinos de 25 a 55 L/dia (CZERKAWSKI, 1969; HOLTER E YOUNG, 1992; McALLISTER et al., 1996), o que corresponde a emissões anuais de 39,1 a 109,5 kg e de 6,5 a 14,4 kg, respectivamente. A Índia e o Brasil lideram o ranking mundial de emissão total de metano entérico, com 14,5 e 10,3 Tg de CH4/ ano, respectivamente. Quando é considerada apenas a emissão por bovinos, o Brasil é apontado como o maior emissor (9,6 Tg de CH4/ ano), seguido da Índia (8,6 Tg de CH4/ano) e dos Estados Unidos da América (5,1 Tg de CH4/ano) (THORPE, 2009). Segundo resultados preliminares do Segundo Inventário Nacional de Emissões de GEE (MCT, 2009), no ano de 2005 a agropecuária foi responsável por 22% do total das emissões de metano no Brasil.
Além de ser caracterizado como um importante GEE, responsável por 15% do aquecimento global, o metano de origem entérica tem relação direta com a eficiência da fermentação ruminal em virtude da perda de carbono e, consequentemente, perda de energia, influenciando o desempenho animal (COTTON E PIELKE, 1995). O conhecimento dos mecanismos de síntese de metano e os fatores que afetam sua produção são importantes. O desafio no sistema produtivo de ruminantes é desenvolver dietas e estratégias de manejo que minimizem a produção relativa de metano (metano/kg de leite, carne ou lã), possibilitando maior eficiência produtiva e redução da contribuição negativa da pecuária para o aquecimento global.
A mídia tem rotulado os bovinos como grandes vilões das mudanças climáticas, sendo que, na maioria das vezes, essas críticas não apresentam fundamentação técnico-científica. É urgente a necessidade de desenvolver e validar metodologias acuradas de mensuração da emissão de metano e gerar bancos de dados específicos para os sistemas de produção de cada região (país ou bioma), conforme relatado no primeiro inventário nacional de emissões de GEE de origem antrópica (GRAINGER et al., 2007; LIMA et al., 2006). A exploração equivocada da mídia sobre o assunto pode ser, futuramente, um pretexto para a criação de barreiras não tarifárias à exportação de produtos pecuários brasileiros.
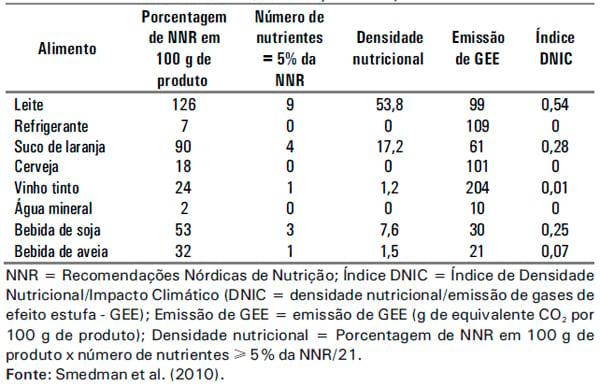
Discussões sobre como reduzir as emissões de GEE têm focado tanto alterações na cadeia de produção e abastecimento de alimentos, como na demanda, por meio de mudanças significativas nos padrões de consumo. Medidas políticas que levam a reduções radicais no consumo de alimentos de origem animal têm sido propostas como meio de reduzir as emissões globais de GEE. Entretanto, a avaliação do impacto climático da produção de diferentes alimentos deve levar em consideração o valor nutricional dos mesmos. Smedman et al. (2010) utilizaram uma unidade funcional, que combina a densidade de nutrientes do alimento com a emissão de GEE na produção dos mesmos, denominada índice de Densidade Nutricional/ Impacto climático (DNIC). Os autores compararam a emissão de GEE geradas para a produção de leite, refrigerantes, suco de laranja, cerveja, vinho, água mineral gasosa e bebidas de soja e aveia. Para a produção de leite foram gerados para cada 100 g do produto, 99 g de equivalente CO2, um dos valores mais elevados quando comparado às demais bebidas. Entretanto, quando a comparação foi realizada levando-se em consideração o DNIC (densidade de nutrientes/emissão de GEE), o leite apresentou vantagem em relação aos demais alimentos, devido ao seu alto valor nutricional (Tabela 1). Esse resultado representa argumento convincente de embate à mídia, que muitas vezes incentiva a redução no consumo de produtos de origem animal como forma de diminuir os impactos ambientais.
Tabela 1. Densidade nutricional, em relação ao impacto climático.
É provável que a agropecuária seja cada vez mais afetada pelas imposições de limitações nas emissões de carbono e pela legislação ambiental. A tendência ou obrigação legal de mitigar as emissões de GEE influenciará diretamente a necessidade de aumento da eficiência zootécnica nos sistemas pecuários, atrelado ao manejo nutricional dos animais a ser adotado. A melhoria das práticas alimentares pode reduzir a emissão de metano por quilograma de alimento ingerido ou por quilograma de produto. Agentes específicos e aditivos dietéticos têm sido propostos como alternativas para a redução das emissões de metano. O desenvolvimento de estratégias de mitigação e a viabilidade da aplicação prática dessas estratégias são áreas atuais de pesquisa em todo o mundo (THORNTON, 2010).
Objetivou-se com o presente artigo abordar diversos aspectos relacionados às emissões de metano entérico pelos ruminantes, bem como descrever as principais metodologias de avaliação e estratégias de mitigação.
Formação do metano entérico e seu papel no ecossistema ruminal
A fermentação dos componentes dietéticos pela microbiota ruminal resulta na formação de ácidos graxos voláteis (AGVs), usados pelo ruminante como fonte de energia, e produção de gases (CO2 e CH4), eliminados por meio da eructação (MARTIN et al., 2009a).
A fermentação em ruminantes envolve processo oxidativo, gerador de cofatores reduzidos (NADH, NADPH e FADH). Para que o processo fermentativo não seja paralisado, esses cofatores são então re-oxidados (NAD+, NADP+ e FAD+) por meio de reações de desidrogenação, liberando hidrogênio no rúmen.
Como processo aceptor de elétrons, a metanogênese remove continuamente o gás Hidrogênio (H2) do meio. Dessa forma, a formação de metano é essencial para o ótimo desempenho do ecossistema ruminal, porque evita o acúmulo de H2 no rúmen, o que poderia levar à inibição da atividade desidrogenase, envolvida na re-oxidação dos cofatores reduzidos. A remoção eficiente do H2 do rúmen contribui para o aumento da taxa de fermentação pela eliminação do seu efeito inibitório na degradação microbiana de materiais vegetais (WOLIN, 1979; McALLISTER E NEWBOLD, 2008).
O metano entérico é derivado da atividade das Archaea metanogênicas, um grupo microbiano distinto das Eukarya (protozoários e fungos) e Bacteria, possuindo cofatores (coenzima M, F420 e F430) e lipídeos (ésteres de isopranil glicerol) únicos. Diante da função central do H2 no metabolismo, as metanogênicas são extremamente importantes para o funcionamento do rúmen e nutrição animal, apesar de responderem por pequena parte da biomassa microbiana ruminal (JANSSEN E KIRS, 2008).
Como as Archaea metanogênicas são responsáveis pela produção de metano nos ruminantes, considerável esforço de pesquisa tem sido direcionado para caracterizá-las (ATTWOOD et al., 2008). A identificação de todo o espectro e diversidade desses microrganismos é condição sine qua non para o desenvolvimento de estratégias de mitigação da emissão de metano entérico. O sequenciamento de seus genomas fornecerá importantes informações que indicarão os alvos mais adequados para estratégias mitigadoras (BUDDLE et al., 2010).
Na complexa comunidade microbiana do trato gastrintestinal, muitos outros microrganismos exercem importante influência na produção de metano, seja por proporcionar ambiente adequado para a sobrevivência das metanogênicas ou por produzir substratos utilizados pelas mesmas. As vias metabólicas envolvidas na formação de hidrogênio, bem como as relações interespécies da população metanogênica com os demais microrganismos do ecossistema ruminal, são importantes fatores que devem ser considerados no desenvolvimento de estratégias para o controle da emissão de metano por ruminantes.
O H2 produzido durante a fermentação microbiana do alimento é usado como fonte de energia pelas Archaea metanogênicas, que produzem metano. O formato também pode ser utilizado pelas metanogênicas, mas é menos importante como precursor do metano do que o H2, contribuindo com aproximadamente 18% da produção (HUNGATE et al., 1970). O ciclo de formação do metano pelas Archaea metanogênicas a partir do CO2 envolve a captação de quatro moléculas de H2:
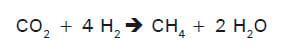
As archaebactérias são também capazes de utilizar o H2 na redução de moléculas de acetato, metilamina e metanol para a produção de metano (MOSS, 1993; WOLIN et al., 1997).
Os diferentes produtos formados durante a fermentação ruminal (AGVs) não são equivalentes em termos de liberação de H2. Portanto, a quantidade de H2 livre liberado no rúmen depende da concentração e proporções relativas de acetato, propionato e butirato produzidos (OWENS E GOETSCH, 1988; EUN et al., 2004; MARTIN et al., 2009a). Existem modelos matemáticos quantitativos que propõem cálculos estequiométricos da fermentação para balancear a formação de H2, ácidos graxos voláteis e outros produtos, na tentativa de predizer a formação de metano (BANNINK et al., 2006; ELLIS et al., 2008a).
A produção de acetato e butirato, predominante durante a fermentação de carboidratos fibrosos, resulta em liberação líquida de H2 e favorece a metanogênese. Já a formação de propionato é uma via competitiva de utilização de H2 no rúmen, reduzindo a disponibilidade de substrato para a metanogênese. Assim, a produção de metano, depende do balanço de H2 no rúmen, sendo influenciada pelas taxas de produção de acetato e propionato (HEGARTY, 2001), como descrito por Van Soest (1994):
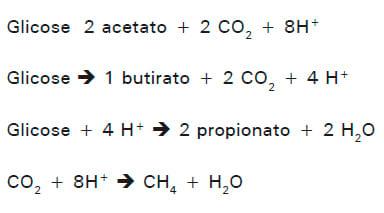
De acordo com a estequiometria de produção dos AGVs, percebe-se o efeito negativo do propionato sobre a metanogênese, devido à competição pelo mesmo substrato (MOSS et al., 2000), gerando uma relação expressa como:
CH4 = 0,45 x Acetato – 0,275 x Propionato + 0,40 x Butirato
Hidrogênio e a produção de metano entérico
A quantidade absoluta de CH4 formado por animais consumindo diferentes dietas está relacionada a características inerentes ao alimento, incluindo a natureza e quantidade de alimento, a extensão de sua degradação, e a quantidade de H2 formado (JOHNSON E JOHNSON, 1995; PELCHEN E PETERS, 1998; JANSSEN, 2010). De forma geral, o aumento da taxa de digestão do alimento está associado ao aumento da taxa de passagem no rúmen. Também está associado a uma menor produção de CH4 por unidade de alimento digerido no rúmen (JANSSEN, 2010). O pool total de H2 no rúmen é pequeno, e a concentração de H2 dissolvido é, aproximadamente, de 0,1-50 μM, que é 0,014 a 6,8% da sua máxima solubilidade a 39ºC e pressão de 1 atm (JANSSEN, 2010).
A pressão parcial de H2 (ou concentração de H2 dissolvido) tem efeito bem estabelecido sobre as vias de fermentação que utilizam ou produzem esse gás. Em coculturas com microrganismos utilizadores de H2, os microrganismos produtores de H2 geram maior quantidade desse gás por unidade de carboidrato fermentado se a pressão parcial de H2 é baixa (WOLIN, 1976; REES et al., 1995; MORVAN et al., 1996). Em altas concentrações de H2, a formação de mais H2 pode tornar-se termodinamicamente desfavorável, ou a mudança de energia livre da transformação do substrato através de vias de produção de H2 pode ser menos favorável do que a transformação em outros produtos (JANSSEN, 2010).
Heijnen e van Dijken (1992) e Heijnen et al. (1992) propuseram uma descrição termodinâmica de produção de biomassa a partir de substratos (fonte de energia), baseada no Modelo da Dissipação de Energia de Gibbs, ou seja, mudança de energia livre, ΔGT. O modelo proposto por Gibbs sugere que a quantidade de biomassa formada a partir de um substrato é determinada pela ΔGT, ou seja, a mudança de energia livre entre reagentes e produtos. Uma dada população de células, metabolizando um substrato sob condições com determinada ΔGT, irá produzir mais biomassa (mais células) do que outra população metabolizando o mesmo substrato por meio de uma via com ΔGT menos negativa (ou seja, com menor liberação de energia livre).
Westerhoff e van Dam (1987) propuseram que a taxa de metabolismo está relacionada à mudança de energia livre da transformação do substrato pelo microrganismo. Ambas, a taxa de metabolismo do substrato para produzir energia metabolicamente útil e a quantidade de biomassa capaz de ser formada a partir do substrato determinarão a taxa de crescimento (μ) de um microrganismo (ou seja, a taxa com que novas células são formadas). Isso implica que μ está relacionada ao ΔGT para um dado substrato, e aquelas espécies metabolizando um substrato por uma via que resulta em ΔGT mais negativo, irão ultrapassar as outras utilizando via com ΔGT menos negativo (JANSSEN, 2010). Portanto, pode-se esperar que aquelas espécies que utilizarem vias de fermentação com ΔGT mais negativo irão dominar a comunidade sob quaisquer condições. Essa é a base pela qual metanogênicas competem com as acetogênicas pelo H2 (CONRAD et al., 1986; CORD-RUWISCH et al., 1988; UNGERFELD E KOHN, 2006).
As mudanças de ΔGT das diferentes vias de fermentação da glicose são influenciadas pela concentração de H2 no rúmen, sendo que algumas vias são mais afetadas do que outras. O ΔGT é mais negativo (ou seja, mais energia é liberada e disponibilizada para acoplar a processos relacionados ao crescimento) para vias de formação de H2 em condições de baixas comparadas às de altas concentrações desse gás. Em contraste, vias que não resultam em formação de H2 não são influenciadas por sua concentração no meio. Essa análise sugere que a formação de butirato + H2 ou acetato + butirato + H2 serão as vias mais favoráveis em baixas concentrações de H2, enquanto que a formação de acetato + propionato deve ser favorecida por altas concentrações desse gás. A consequência do efeito da concentração de H2 sobre a termodinâmica da fermentação é que, sob condição em que a concentração desse gás no rúmen é elevada, sua produção torna-se desfavorável e as vias de formação de propionato são as mais favorecidas (JANSSEN, 2010).
Proteínas são também fermentadas no rúmen, com consequente produção de metano. Entretanto, mudanças na concentração de H2 não favorecem vias alternativas de fermentação de aminoácidos da mesma forma que favorecem diferentes vias de fermentação de glicose. Portanto, variações nas condições ruminais que têm algum efeito sobre a metanogênese e concentrações de H2 não terão grande impacto sobre a formação de H2 e metano a partir da fermentação de proteína, como terão a partir da fermentação de carboidratos (JANSSEN, 2010).
As relações entre a concentração de H2 e a cinética de crescimento das Archaea metanogênicas, e os efeitos do H2 na termodinâmica de fermentação são prontamente compreendidas quando aplicadas a um sistema homogêneo. Entretanto, o rúmen é um sistema dinâmico e heterogêneo (CZERKAWSKI, 1986), apresentando grande número de pequenos microambientes transitórios. Em cada microambiente, os microrganismos ruminais estão degradando alimento, produzindo H2 e convertendo-o em CH4. Em alguns, a produção e utilização de H2 estarão estreitamente acopladas, e a concentração desse gás será baixa, devido à atividade das metanogênicas. Em outros, a rápida fermentação dos nutrientes alimentares resultará em concentração elevada de H2. Em todo rúmen, a extensão de tempo com que cada um desses pequenos sistemas existe e a relativa abundância de microambientes com alta versus baixa concentração de H2 determinarão o balanço entre as vias que produzem diferentes quantidades desse gás e a seleção de espécies de microrganismos que crescem melhor em baixas ou altas concentrações do mesmo. Assim, o grau com que mudanças nos microambientes ocorrem será modelado pelo tempo que os microrganismos têm para alterar seus padrões de fermentação, bem como aquele que a microbiota leva para ser modificada. O balanço líquido da taxa de crescimento das metanogênicas no rúmen controla a concentração média de H2 nesse ambiente. A soma de todas essas mudanças determina a quantidade líquida de H2 formada e, portanto, a quantidade líquida de CH4 produzida por unidade de alimento consumido (JANSSEN, 2010).
Transferência de H2 interespécies
As Archaea metanogênicas estão na base da cadeia trófica microbiana e utilizam como substratos os produtos finais da fermentação. A captura do H2 produzido por uma determinada espécie ruminal por outra é normalmente referido como transferência de H2 interespécies (WOLIN et al., 1997), sendo um processo que, em muitos casos, envolve uma relação simbiótica entre dois microrganismos.
Os microrganismos fibrolíticos exercem papel central no ecossistema ruminal. De fato, eles estão no primeiro nível da cadeia trófica microbiana, transformando os polissacarídeos da parede celular dos alimentos em AGVs, CO2 e H2. A maior parte dos microrganismos fibrolíticos produz H2 como principal produto final da fermentação que, por sua vez, sob condições fisiológicas normais, é rapidamente utilizado pelas metanogênicas (MORGAVI et al., 2010).
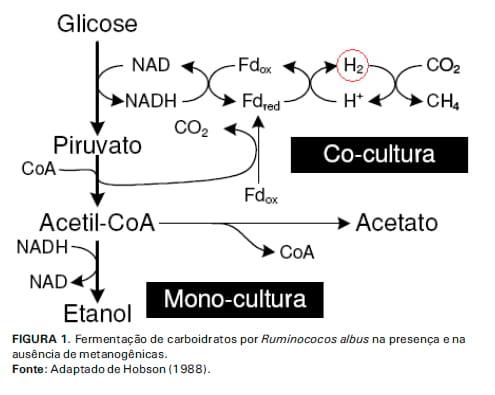
O metabolismo de duas espécies ruminais importantes na degradação de carboidratos fibrosos (Ruminococcus albus e R. flavefasciens) é influenciado pela pressão parcial de H2 no rúmen e, portanto, pela atividade das metanogênicas. O sistema de transferência de H2 interespécies acopla a reação oxidativa da espécie celulolítica R. albus com a reação redutora da comunidade Archaea metanogênica. (Figura 1). Em monocultura, as bactérias R. albus produzem etanol, acetato, H2 e CO2. Quando em cocultura com metanogênicas, o NADH é utilizado para reduzir prótons a H2, dando origem a acetato e CH4 como produtos finais. Dessa forma, a formação de etanol é evitada, levando à maior produção de ATP por unidade de hexose fermentada (WOLIN E MILLER, 1988).
Os efeitos da diversidade de espécies das comunidades microbianas fibrolíticas (produtores e não produtores de H2) sobre a produção de metano têm sido investigado. A incubação in vitro do conteúdo ruminal de cordeiros gnotobióticos indicou maior produção de metano pelo inóculo contendo microrganismos fibrolíticos produtores de H 2 (Ruminococci ou fungos) do que por aquele contendo microrganismos fibrolítico s não produtores de H2 (Fibrobacter) (CHAUCHERYAS-DURAND et al., 2008). Morvan et al. (1996) observaram correlação positiva entre o número de bactérias fibrolíticas e o de microrganismos metanogênicos no rúmen de vários animais, incluindo bovinos, ovinos, lhamas e veados, e no ceco de equinos. Tal correlação foi encontrada porque as principais espécies fibrolíticas, tais como R. albus e R. flavefasciens, produzem H2 como principal produto da fermentação. Em contraste, os mesmos autores relataram que no rúmen de búfalos, o número de microrganimos metanogênicos foi comparativamente menor do que o de bactérias fibrolíticas. Tal fato pode ter ocorrido porque nestes animais as bactérias Fibrobacter, as quais não produzem H2, eram os microrganismos fibrolíticos predominantes no rúmen (MORVAN et al., 1994).
No rúmen, as Archaea metanogênicas têm sido encontradas intimamente associadas aos protozoários, e essa adesão parece ser reversível (STUMM et al., 1982; TOKURA et al., 1997). Os protozoários ciliados do rúmen são metabolicamente muito ativos, capazes de influenciar a fermentação de alimentos e outras populações microbianas ruminais e, consequentemente, afetar a quantidade e proporção de produtos finais da fermentação ruminal, incluindo metano (WILLIAMS E COLEMAN, 1992; EUGÈNE et al., 2004). Mais de 37% do metano derivado do rúmen pode ser produzido por metanogênicas associadas a protozoários (FINLAY et al., 1994; NEWBOLD et al., 1995).
Protozoários engolfam matéria orgânica, particularmente bactérias, dentro de vacúolos digestivos, onde a hidrólise e fermentação acontecem. Os principais AGVs produzidos são acetato e butirato (WILLIAMS E COLEMAN, 1992). Os protozoários contribuem para a metanogênese por meio do fornecimento de H2 produzido durante a fermentação dos carboidratos, servindo de hospedeiros para cerca de 30% das Archaea metanogênicas (JOUANY, 1996), e protegendo-as da toxicidade do O2 (MORGAVI et al., 2010). O H2 é produzido em grande quantidade pelos protozoários em organelas equivalentes às mitocôndrias dos eucariotas aeróbicos: o hidrogenossoma. Esse H2 é utilizado pelas metanogênicas que estão dentro ou em estreita associação com as células protozoárias (STUMM et al., 1982; FINLAY et al., 1994).
A associação somática das metanogênicas com protozoários ciliados representa típica relação simbiótica de transferência de H2 interespécies, em que ambos são favorecidos. As metanogênicas, por utilizarem o H2 produzido pelos ciliados, favorecem a manutenção de ambiente ruminal adequado ao desenvolvimento desses microrganismos (PEDREIRA et al., 2005), permitindo que a fermentação da matéria orgânica seja direcionada mais para a produção de acetato e CO2 em detrimento de butirato e lactato, o que resulta em maior eficiência na produção de ATP pelo protozoá rio hospedeiro (MORGAVI et al., 2010). A extensão da associação entre metanogênicas e protozoários é influenciada pela dieta e pelo tempo após alimentação (TOKURA et al., 1997).
***O Trabalho foi originalmente publicado pela Embrapa Gado de Leite/ Maio,2011
Tópicos relacionados
Autores:

Mostrar mais
Junte-se para comentar.
Uma vez que se junte ao Engormix, você poderá participar de todos os conteúdos e fóruns.
* Dados obrigatórios
Quer comentar sobre outro tema? Crie uma nova publicação para dialogar com especialistas da comunidade.
Criar uma publicação