INTRODUÇÃO
O triptofano desempenha vários papéis no metabolismo das aves, como precursor da niacina e serotonina. Pelo fato de ser importante no sistema imunológico e ter nos seus metabólicos os principais produtos envolvidos na regulação do consumo e do estresse, é fundamental o estabelecimento do nível correto deste aminoácido nas rações. A suplementação de L-Triptofano contribui para uma melhor utilização dos outros aminoácidos e da ração em geral. O L-Triptofano 98% é produzido por fermentação de matéria-prima de origem agrícola, como o melaço de beterraba ou o hidrolisado de amido. Consiste em cristais branco a amarelo claro com 98% de triptofano e 100% de digestibilidade (AJINOMOTO, 2010).
Segundo alguns autores, como Peganova et al. (2003) e Deponti et al. (2007), o triptofano é considerado como o terceiro aminoácido limitante nas rações de galinhas poedeiras à base de milho e farelo de soja. Além disso, o triptofano é precursor da síntese de serotonina e melatonina, o que, sem dúvida alguma facilita em muito o manejo diário das aves e, de certa forma, contribui para uma produção de ovos mais eficiente, haja vista o número de ovos com avarias que é reduzido nestas situações. Sua deficiência provoca uma redução no consumo de ração (Peganova & Eder, 2002) e sua exigência pode ser influenciada por aminoácidos neutros de cadeia longa presentes nas dietas de poedeiras, como a isoleucina, valina, leucina, fenilalanina, tirosina, metionina e histidina, e ainda mais pela proteína da dieta.
Diferente das recomendações de lisina, o triptofano é mais avaliado com base na relação com a lisina, ou seja, além dos níveis individuais do aminoácido, a relação com a lisina passa a ser, atualmente um dos maiores aliados dos pesquisadores em determinar o melhor nível do aminoácido. Assim, para a fase de postura, as recomendações são as mais diversas. Como forma de exemplo o NRC. (1994), o CVB (1996), Coon & Zhang. (1999), Rostagno et al. (2000), Rostagno et al. (2005), Rostagno et al. (2011), Leeson & Summers. (2005) e Brengendahl (2008) recomendam que as dietas desta aves, sejam elas leves ou semipesadas, tenham uma relação triptofano digestível: lisina digestível em 23, 19, 20, 25, 23, 23, 21 e 22, respectivamente.
Assim, este estudo teve como objetivo determinar a exigência de triptofano digestível para poedeiras leves alimentadas com rações com níveis crescentes de triptofano sob relação variável e constante.
MATERIAL E MÉTODOS
O estudo foi realizado no Módulo de Pesquisas com Aves do CCA/UFPB, Areia/PB. A cidade de Areia está inserida na microrregião do Brejo Paraibano (6º 58' 12'' de latitude Sul e 35º 42' 15'' de longitude Oeste), com altitude média de 619 m (GONDIM e FERNANDES, 1980). De acordo com a classificação de Köppen, o clima é tropical semi-úmido do tipo As', isto é, clima quente e úmido que se caracteriza por apresentar estação chuvosa no período outono-inverno, com precipitação pluviométrica média anual girando em torno de 1500 mm, temperatura média anual entre 22 a 30ºC e umidade relativa do ar elevada (75 a 87%). Segundo a classificação bioclimática de Gäussen, nesta região predomina o bioclima 3 dth (nordestino sub-seco), com período seco variando de um a três meses no ano (BRASIL, 1972).
Foram desenvolvidos dois experimentos seqüenciais, o primeiro com rações que continham níveis crescentes de triptofano digestível, alterando-se a relação triptofano digestível: lisina digestível e, com base nos resultados, realizou-se um segundo experimento, com o objetivo de determinar a exigência de triptofano digestível com relação aminoácidos: lisina constante.
Experimento 1 – Níveis crescentes de triptofano digestível, relação variável
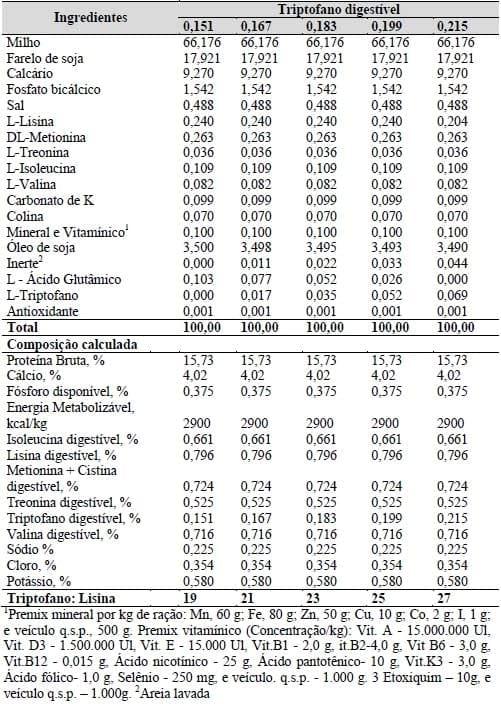
Um total de 280 galinhas poedeiras leves da linhagem Dekalb White com 29 semanas de idade, distribuídas em um delineamento inteiramente ao acaso, com cinco tratamentos, 14 repetições com 4 aves cada foram utilizadas. O experimento foi dividido em 5 fases de 28 dias cada, totalizando 140 dias de avaliação. Os tratamentos consistiram em uma ração basal, formulada à base de milho e farelo de soja, suplementada com os aminoácidos sintéticos L-Lisina, DL-Metionina, L-Treonina, L-Isoleucina e L-Valina de forma a atender as exigências nutricionais para poedeiras, exceto para triptofano digestível. A dieta basal foi suplementada com 0,00; 0,017; 0,035; 0,052 e 0,069% de L-Triptofano em substituição ao amido de milho com o objetivo de alcançar os níveis de 0,151; 0,167; 0,183; 0,199 e 0,215% de triptofano digestível na ração (Tabela 17).
As relações aminoácidos: lisina (em aminoácidos digestíveis) atenderam as recomendações de Rostagno et al. (2005), exceto as triptofano digestível: lisina digestível, que foi de 19, 21, 23, 25 e 27 para cada tratamento. O carbonato de potássio foi acrescido na ração para atender os níveis de potássio exigidos, em função da redução da quantidade de farelo de soja, que foi limitada pela necessidade de se causar a deficiência de triptofano na dieta basal. O L - Ácido Glutâmico foi utilizado para tornar as dietas isoprotéicas. O nível dietético do óleo de soja e do inerte foi equilibrado em substituição ao L - Ácido Glutâmico que foi reduzido com o aumento dos níveis de L-Triptofano nas rações experimentais, de forma a tornar as dietas, além de isoprotéicas, isoenergéticas.
Tabela 17. Composição alimentar e química das rações experimentais.
Experimento 2 – Níveis crescentes de triptofano digestível, relação constante
Foram utilizadas 280 poedeiras leves da linhagem Dekalb White com 29 semanas de idade, distribuídas em um delineamento experimental inteiramente casualizado, com 5 tratamentos, 14 repetições e 4 aves por unidade experimental. . O experimento foi dividido em 5 fases de 28 dias cada, totalizando 140 dias de avaliação.
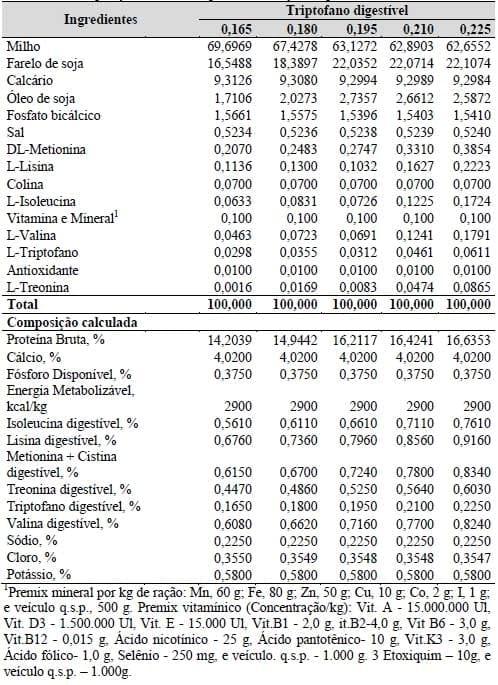
Os tratamentos consistiram em uma ração formulada à base de milho e farelo de soja, suplementada com os aminoácidos industriais L-Lisina, DL-Metionina, L-Triptofano, L-Isoleucina e L-Valina de forma a atender as exigências nutricionais para poedeiras leves em postura. As rações tinham a mesma relação aminoácidos: lisina de acordo com as recomendações sugeridas por Rostagno et al. (2005), exceto a triptofano digestível: lisina digestível que foi mantida constante em 24,5%. As relações aminoácidos: lisina digestível foram 91, 66, 90, 83 e 24,5%, para metionina+cistina, treonina, valina, isoleucina e triptofano digestíveis, respectivamente, conforme apresentadas na Tabela 18.
Variáveis avaliadas
As variáveis avaliadas foram: consumo de ração (g/ave/dia), produção (%), peso (g), massa de ovo (g/ave/dia), conversão por massa (kg/kg) e por dúzia de ovo (kg/dz), peso (g) e porcentagem (%) de gema, de albúmen e de casca, espessura da casca (mm), unidade Haugh, gravidade específica.
O período de avaliação da produção de ovos foi dividido em cinco períodos de 28 dias cada. Ao final de cada período foram coletadas as sobras das rações de cada parcela para o cálculo do consumo de ração. A coleta dos ovos foi realizada duas vezes ao dia (10:00 e 16:00 h), sendo anotados em ficha de freqüência de postura e a mortalidade.
Tabela 18. Composição alimentar e química das rações experimentais.
A produção dos ovos em porcentagem foi calculada dividindo-se a quantidade de ovos totalizados por parcela pelo número de aves, corrigindo pela mortalidade. Os ovos dos últimos três dias de cada período foram pesados individualmente para a obtenção do peso médio dos ovos. Os cálculos da massa de ovo foram realizados pelo produto da produção de ovos e do peso médio dos ovos por parcela. A conversão alimentar por massa de ovo foi calculada através da relação entre o consumo de ração e massa de ovo produzida. A conversão por dúzia de ovos foi calculada pela relação entre o consumo de ração dividida pela produção, sendo esse resultado multiplicado por doze.
Ao final de cada período, foram selecionados quatro ovos por parcela para determinação do peso e porcentagem de gema, de albúmen e de casca, após separação manual destes componentes, onde as cascas foram colocadas em estufa a 105°C por quatro horas. A porcentagem de cada um dos componentes do ovo foi obtida dividindo-se o peso do componente pelo peso do ovo, em seguida, multiplicando o resultado por 100. A espessura da casca foi medida com o auxílio de um micrômetro digital com precisão de 0,1 mm em três pontos na linha mediana do ovo, com os quais foi calculada a média aritmética.
A cada final de período experimental foram selecionadas amostras representativas de dois ovos por parcela onde foram feitas imersões dos ovos em diferentes soluções salinas com os devidos ajustes para um volume de 25 litros de água com densidades que variavam de 1,060 a 1,100 com intervalo de 0,0025 g/cm3. Os ovos eram colocados nos baldes com as soluções, da menor para a maior densidade e eram retirados ao flutuarem, sendo anotados os valores respectivos das densidades correspondentes às soluções dos recipientes. Antes de cada avaliação, as densidades eram conferidas com densímetro de petróleo.
Nas análises histológicas, a coleta do material foi realizada através de fragmentos do sistema digestório (duodeno e fígado) e reprodutor (magno e útero) de 10 animais por tratamento. Os fragmentos foram imersos em fixador metacarn (60% metanol, 30% clorofórmio e 10% ácido acético) por 12 horas, sendo transferidos em seguida para álcool 70%.
Para a microscopia óptica foi feita a inclusão dos fragmentos em paraplast. Foram realizados cortes seriados dos fragmentos com 5 μm de espessura. As seguintes colorações histológicas foram realizadas para a descrição histológica: hematoxilina e eosina, periodic acid Schiff (PAS) e tricômio de Masson. As fotomicrografias foram capturadas com o auxílio de microcâmera acoplada ao microscópio Olympus BX-51 e as imagens digitalizadas no software KS 400.3 (Zeiss).
Os resultados foram submetidos as analises de variância e regressão utilizando-se o programa SAEG (Universidade Federal de Viçosa, 2000).
RESULTADOS
Experimento 1 – Níveis crescentes de triptofano digestível, relação variável
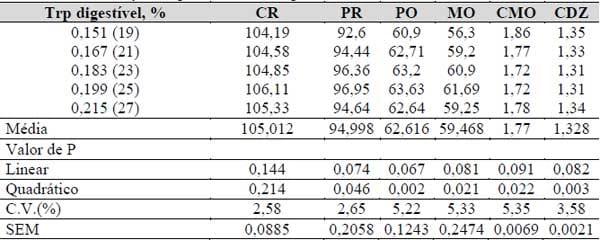
A variação dos níveis de triptofano digestível influenciou o desempenho das galinhas poedeiras leves, para todas as variáveis avaliadas, exceto para o consumo de ração (Tabela 19), apresentando efeito significativo, com comportamento quadrático (P<0,01).
Tabela 19. Consumo de ração (CR, g), produção (PR, %), peso (PO, g), massa (MO, g), conversão em massa (CMO, g/g) e em dúzia de ovos (CDZ, g/g) em função das relações triptofano: lisina digestíveis.
A produção de ovos apresentou um melhor índice com uma relação triptofano digestível: lisina digestível em 23,98%. Já o peso, massa, conversão em massa e em dúzia de ovos, tiveram recomendações semelhantes, ou seja, 24,07; 24,11; 24,58 e 25,25, respectivamente.
Tabela 20. Equações polinomiais das variáveis influenciadas estatisticamente pelas relações triptofano: lisina digestíveis.
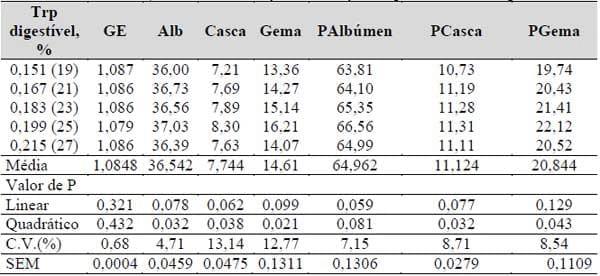
Com exceção da gravidade específica, os níveis de triptofano das rações influenciaram (P<0,05), os pesos absolutos do albúmen, casca e gema, e os pesos relativos da casca e gema. Embora os dados brutos sejam reais, os pesos relativos parecem ter maior representatividade na avaliação, devido especialmente, aos melhores coeficientes de variação obtidos, conforme Tabela 21.
Tabela 21. Gravidade específica (GE, g/cm3), pesos absolutos e relativos do albúmen (Alb, g; PAlbúmen, %), casca (Casca, g; PCasca, %) e gema (Gema, g; PGema, %) dos ovos em função das relações triptofano: lisina digestíveis.
As variáveis apresentadas na Tabela 21 com efeito significativo tiveram um comportamento quadrático, de modo que na Tabela 22, são apresentadas as equações polinomiais e os respectivos dados para os pesos absolutos do albúmen, casca e gema, e pesos relativos da casca e gema, o que representa níveis de 0,192, 0,193, 0,190, 0,196, 0,192% de triptofano digestível, respectivamente.
Tabela 22. Equações polinomiais das variáveis influenciadas estatisticamente pelas relações triptofano: lisina digestíveis.

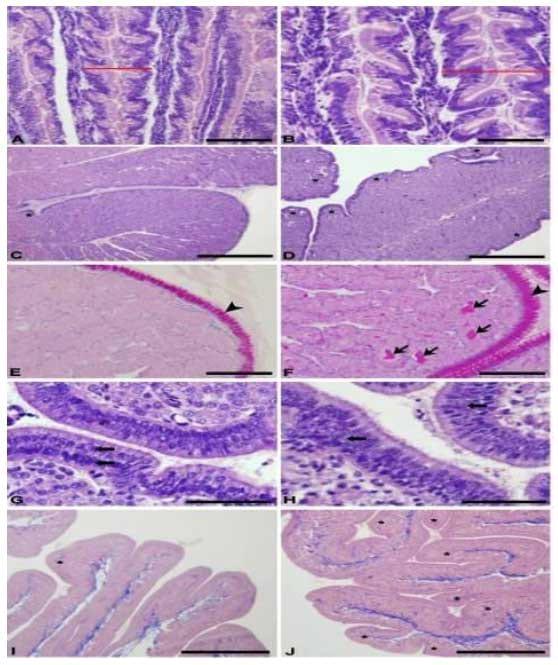
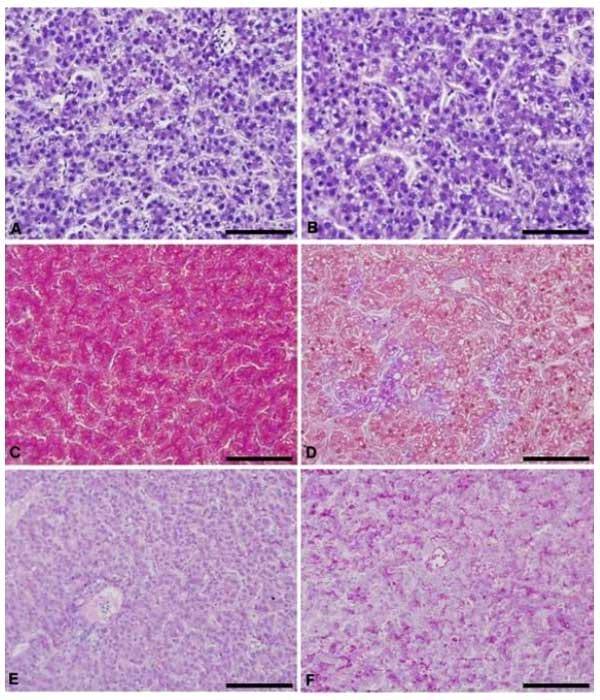
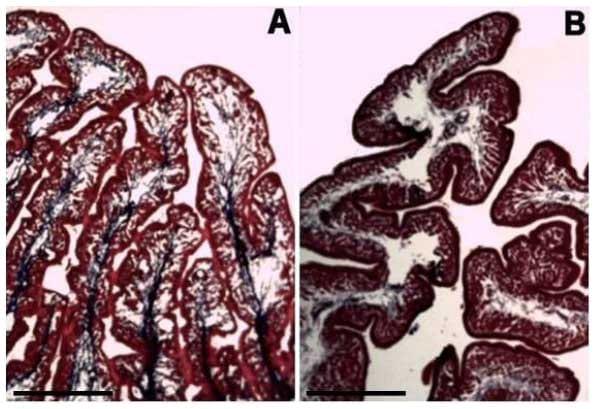
Histologicamente foi verificado nas poedeiras alterações em alguns órgãos dos sistemas reprodutor e digestório em decorrência das rações contendo maior nível de triptofano digestível, especialmente naquelas com níveis acima de 0,183% de triptofano digestível. As vilosidades intestinais das poedeiras alimentadas com rações com níveis acima de 0,183% de triptofano digestível se apresentaram mais largas e possuíam mais ramificações (Figura 9A, B). Não foi observado alteração na quantidade nem na produção de células caliciformes intestinais. As dobras do magno, nesses mesmos tratamentos, se apresentaram mais ramificadas e com glândulas mais ativas (Figura 9C, D). O epitélio dessas dobras mostrou uma maior produção de muco (Figura 9E, F). O útero das poedeiras nesses tratamentos apresentou mais dobras secundárias (Figura 9I, J) e o epitélio apresentou hiperplasia (Figura 9G, H).
Figura 9. Fotomicrografia de órgãos do sistema digestório e reprodutor de poedeiras submetidas à rações com diferentes níveis de triptofano digestível. A) Intestino de poedeira alimentada com ração contendo 0,151% de triptofano digestível. B) Intestino de poedeira alimentada com ração contendo níveis maiores que 0,183% de triptofano digestível. C) Magno de poedeira alimentada com ração contendo 0,151% de triptofano digestível. D) Magno de poedeira alimentada com ração contendo níveis maiores que 0,183% de triptofano digestível. E) Magno de poedeira alimentada com ração contendo 0,151% de triptofano digestível. F) Magno de poedeira alimentada com ração contendo níveis maiores que 0,183% de triptofano digestível. G) Epitélio do útero de poedeira alimentada com ração contendo 0,151% de triptofano digestível. H) Epitélio do útero de poedeira alimentada com ração contendo níveis maiores que 0,183% de triptofano digestível. I) Dobras do útero de poedeira alimentada com ração contendo 0,151% de triptofano digestível. J) Dobras do útero de poedeira alimentada com ração contendo níveis maiores que 0,183% de triptofano digestível. A, B, C, D, G e H: Hematoxilina-eosina. E e F: Periodic acid Schiff. I e J: Tricômio de Masson. Barra: G e H- 50 μm; A, B, E e F- 100 μm; C, D, I e J- 500 μm.
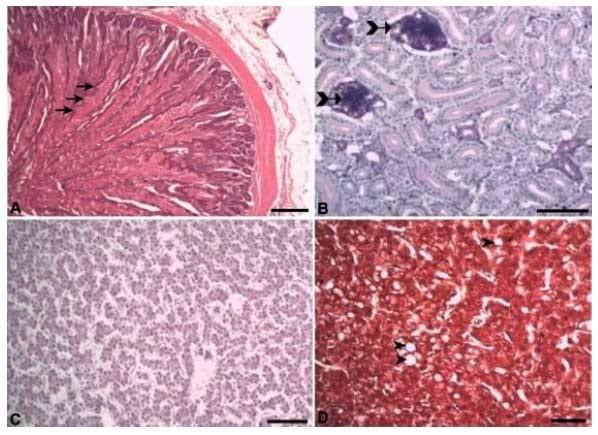
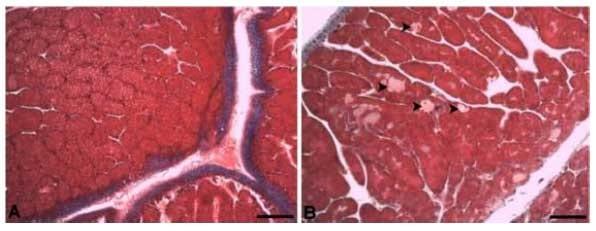
Com o aumento do nível de triptofano digestível nas rações houve também um aumento da esteatose hepática (Figura 10A, B). No tratamento em que as aves foram submetidas à ração com 0,215% de triptofano digestível, ou relação de 27%, fica evidente a esteatose hepática (Figura 10B, D). Nas aves alimentadas com o maior nível de triptofano na ração também foi encontrado pequena quantidade de colágeno parenquimal (Figura 10D). Esse colágeno parenquimal não é encontrado nas aves alimentadas com menor nível de triptofano digestível, ou seja, com 0,151 e 0,167% de triptofano digestível, o que corresponde a uma relação triptofano: lisina digestíveis de 19 e 21%, respectivamente (Figura 10C). No fígado das poedeiras alimentadas com 0,215% de triptofano digestível, relação de 27% com a lisina, houve uma maior positividade para a coloração histológica PAS (Periodic Acid Schiff), denotando maior depósito de glicogênio (Figura 10E, F).
Figura 10. Fotomicrografia do fígado de poedeiras alimentadas com rações contendo níveis crescentes de triptofano digestível. A, C, E) fígado de poedeiras alimentadas com 0,151% de triptofano digestível. B, D e F) fígado de poedeiras alimentadas com 0,215% de triptofano digestível.. A, B) Hematoxilina-eosina. C, D) Tricômio de Masson. E, F) PAS. Barra: 100 μm.
É possível observar que o fígado das aves alimentadas com 0,215% de triptofano digestível apresentou hepatócitos com vacúolos lipídicos citoplasmáticos, indicativo de esteatose (B e D). Nesse grupo também houve o depósito de colágeno parenquimal, representado pela coloração azulada da coloração de Tricômio de Masson (D) e acúmulo de glicogênio hepático representado pela coloração rosa escuro na coloração de Periodic Acid Schiff (PAS) (F).
Experimento 2 – Níveis crescentes de triptofano digestível, relação constante
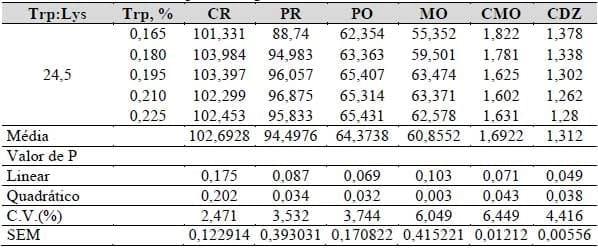
A variação dos níveis de triptofano digestível mantendo-se a relação aminoácidos: lisina constante influenciou o desempenho das galinhas poedeiras leves, conforme apresentado na Tabela 23. O consumo de ração não foi modificado estatisticamente (P>0,05), contudo os demais itens avaliados e apresentados na Tabela 24 apresentaram efeito significativo, com comportamento quadrático (P<0,05 e P<0,01).
Tabela 23. Consumo de ração (CR, g), produção (PR, %), peso (PO, g), massa (MO, g), conversão em massa (CMO, g/g) e em dúzia de ovos (CDZ, g/g) em função dos níveis de triptofano digestível.
Após as derivações das equações polinomiais obtidas pelos efeitos das variáveis, foi obtida as devidas recomendações, ou seja, o ponto de máximo ou de mínino, considerando a variável avaliada. A produção de ovos, o peso , a massa, a conversão em massa e a conversão em dúzia de ovos, com base em suas equações, determinaram como os melhores níveis de triptofano digestível em 0,206, 0,217, 0,209, 0,217 e 0,220%, respectivamente.
Na Tabela 24, são apresentadas as equações polinomiais dos efeitos significativos apresentados anteriormente na Tabela 23.
Tabela 24. Equações polinomiais das variáveis influenciadas estatisticamente pelos níveis de triptofano digestível.
As variáveis que avaliam a qualidade dos ovos não foram influenciadas pelas rações (Tabela 25), tal qual no experimento com relação variável.
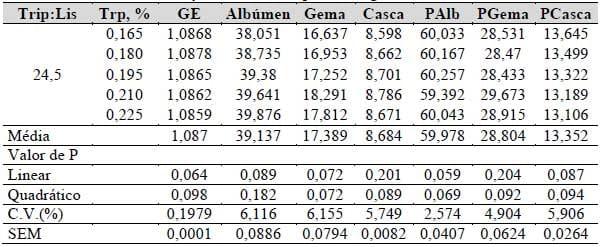
Tabela 25. Gravidade específica (GE, g/cm3), pesos absolutos e relativos do albúmen (Alb, g; Palb, %), casca (Casca, g; Pcasca, %) e gema (Gema, g; Pgema, %) dos ovos em função dos níveis triptofano digestível.
No estudo histológico verificou-se que todos os tratamentos (0,165, 0,180, 0,195, 0,210 e 0,225% de triptofano na ração) conferiram ao intestino vilosidades compridas e bem ramificadas, identificadas com auxílio de ponta de seta (Figura 11A). Os tratamentos também não levaram à lesões renais como demonstra a Figura 11B, onde podemos identificar, com auxílio de uma seta, os glomérulos renais.
O parênquima hepático também se mostrou normal, com manutenção da arquitetura hepática, formada por cordões duplos de hepatócitos, também não houve deposição irregular de glicogênio (Figura 11C). Entretanto, alguns animais do tratamento com 0,165% de triptofano na ração apresentaram moderada esteatose, onde podemos identificar, com auxílio de pontas de seta, os vacúolos citoplasmáticos de lipídeos, entretanto, não foi observada qualquer deposição de colágeno parenquimal (Figura 11D).
Figura 11. Fotomicrografias de poedeiras submetidas à rações com níveis crescentes de triptofano digestível com relação constante. A) Intestino representativo de poedeiras submetidas à dietas com níveis crescentes de triptofano digestível com relação constante. Ramificações identificadas por ponta de seta. Hematoxilina-eosina. Bar: 400 μm. B) Rim representativo de poedeiras submetidas à dietas com níveis crescentes de triptofano digestível com relação constante. As setas mostram os glomérulos renais. Periodic acid Schiff. Bar: 100 μm. C) Fígado representativo de poedeiras submetidas à dietas com níveis crescentes de triptofano digestível com relação constante.. Periodic acid Schiff. Bar: 200 μm. D) Parênquima hepático de um dos animais do tratamento com 0,165% de triptofano. Vacúolos citoplasmátivos lipídicos identificados por pontas de seta. Tricômio de Masson. Bar: 100 μm.
O sistema reprodutor mostrou alterações histológicas nos tratamentos com 0,210 e 0,225% de triptofano, principalmente no grupo com 0,210%. Nesse último grupo, as dobras do magno se apresentaram com maior quantidade de glândulas ativas (Figura 12B) quando comparadas às glândulas dos tratamentos com 0,165, 0,180 e 0,195% de triptofano (Figura 12A).
Figura 12. Fotomicrografias do magno de poedeiras submetidas à dietas com níveis crescentes de triptofano digestível com relação constante. A) Magno de poedeira representando os tratamentos de 0,165%, 0,180% e 0,195% de triptofano. B) Magno de poedeira do tratamento com 0,210% de triptofano. As pontas de seta mostram as glândulas em regeneração com acúmulo de albúmen. Tricômio de Masson. Bar: 50 μm.
Nos tratamentos com 0,210% e 0,225% de triptofano digestível na ração também foram observados dobras uterínicas mais ramificadas e compridas, principalmente no nível de 0,210% (Figura 13).
Figura 13. Fotomicrografias do útero de poedeiras submetidas à dietas com níveis crescentes de triptofano digestível com relação constante. A) Útero de poedeira representativa dos tratamentos com 0,165, 0,180 e 0,195% de triptofano digestível. B) Útero de poedeira do tratamento com 0,210% de triptofano digestível na ração. Tricômio de Masson. Bar: 300 μm.
DISCUSSÃO
Os aminoácidos são estruturas químicas muito similares e, em alguns casos, disputam pelo mesmo sítio de absorção, gerando o que se chama de antagonismo aminoacídico. Neste contexto, Peganova & Eder (2003), comentam que o excesso de isoleucina pode comprometer a passagem de triptofano pela barreira hemato-encefálica, levando à deficiência deste aminoácido no cérebro, o que poderia influenciar na síntese ou na liberação de serotonina. Os autores verificaram também que o aumento dos níveis de triptofano de 0,15 para 0,24% resultou em aumento no consumo de ração, porém sem efeito sobre os níveis plasmáticos de triptofano ou sobre a massa de ovo produzida.
Diferentemente do observado por Peganova & Eder (2003), no experimento com níveis crescentes de triptofano e com relação variável não foi observado efeito sobre o consumo de ração, contudo, houve, mesmo que numericamente, um maior consumo de ração nos tratamentos com maior nível desse aminoácido. A diferença entre os resultados deste com o estudo dos autores supracitados pode advir da manutenção dos níveis protéicos e energéticos das rações experimentais, variando apenas o triptofano, enquanto que no trabalho de Peganova & Eder (2003) tais níveis não foram equilibrados em função da suplementação de L-Triptofano nas rações.
Em uma avaliação do nível ideal de triptofano na ração de poedeiras com 28 semanas de idade, Harms & Russel (2000) utilizaram níveis crescentes (0,12; 0,13; 0,14; 0,15; 0,16; 0,17; 0,18 e 0,20% de triptofano). Em tal estudo, a produção e o peso dos ovos foram aumentados com o crescimento dos níveis de triptofano. Além disto, as aves apresentaram nos níveis mais baixos de triptofano (0,12, 0,13 e 0,14%), uma redução no peso vivo no final do estudo, enquanto que, nos demais tratamentos, com níveis mais elevados do aminoácido, as aves mantiveram ou chegaram até a ganhar peso. De acordo com os dados, o triptofano melhorou significativamente o desempenho das aves avaliadas pelos autores, ao ponto da produção de ovos passar de 66,20 para 92,70% nas aves que consumiram uma ração com 0,12 e 0,17% de triptofano, respectivamente. Além da produção de ovos, o peso dos ovos teve um aumento expressivo, passando de 49,70 para 54,80g quando as aves consumiam uma ração com 0,12 e 0,20% de triptofano, respectivamente. Para esta variável, os autores estimaram através de regressão broken-line uma necessidade dietética de 0,185% de triptofano.
O aumento do triptofano digestível nas rações das aves avaliadas neste estudo melhoraram a produção e a qualidade dos ovos, conforme também observado por Harms & Russel (2000). Embora o consumo de ração não tenha sido influenciado e com a melhoria na produção e peso dos ovos, as variáveis que são obtidas através dessas, como as conversões em massa e em dúzia de ovos foram, seqüencialmente, melhoradas. Já no estudo com a relação aminoácidos:lisina constante, os níveis crescentes dos aminoácidos melhoraram o desempenho das aves, mas não influenciaram a qualidade dos ovos, diferentemente do experimento inicial, o que pode ser explicado pela variação na proteína bruta deste com relação constante, alterando, possivelmente a relação aminoácidos essenciais: não essenciais.
Deponti et al. (2007) com galinhas poedeiras em postura obtiveram resultados semelhantes aos encontrados neste estudo. Os autores verificaram que a produção e a massa de ovos foram negativamente afetadas durante seis semanas, quando poedeiras receberam dietas com baixos níveis de triptofano (0,13%), sendo o desempenho recuperado após uma semana de alimentação com ração contendo 0,21% de triptofano. Nossos resultados também corroboram com os encontrados por Harms & Russell (2000), quando perceberam um pior desempenho das aves com níveis mais baixos de triptofano na ração das aves.
Bregendahl et al. (2008) em uma série de experimentos com poedeiras leves de 28 a 34 semanas determinaram que a relação ideal de triptofano digestível: lisina digestível era de 22, correspondente ao consumo diário de 538 mg de lisina e 120 mg de triptofano. Nesse estudo, na média das recomendações obtidas no experimento 1, desde os dados de desempenho e qualidade externa e interna dos ovos, uma relação triptofano digestível: lisina digestível de 24,5% foi que proporcionou melhores resultados, o que corresponde a 0,195% de triptofano digestível e um consumo diário de 105 mg/ave. Em experimento conduzido por Costa et al. (2010), também com poedeiras leves, com relações triptofano: lisina digestível variando de 19 a 27% demonstrou-se uma resposta quadrática para todas as variáveis avaliadas, exceto consumo de ração, logo os autores concluíram que a relação triptofano: lisina digestível de 24% foi suficiente para melhor desempenho das aves, o que se confirma neste estudo e nas recomendações feitas por Bregendahl et al. (2008).
Com base ainda nas determinações das exigências de triptofano digestível, no experimento com níveis crescentes de aminoácidos e relação aminoácidos: lisina constante, os melhores resultados foram observados quando as aves se alimentavam com níveis entre 0,206 a 0,220% com média entre as determinações de 0,214% de triptofano digestível. Embora esse nível do aminoácido seja mais elevado que o recomendado em trabalhos recentemente publicados, a relação de triptofano: lisina é similar a recomendada por autores supracitados, o que mostra que não basta a relação do aminoácido em relação à lisina, mas o nível em que a ave apresente seu maior potencial de produção.
Alguns dados na literatura apenas relatem os efeitos da suplementação de triptofano nas rações sobre o desempenho zootécnico, os resultados conciliam esses dados com as modificações histológicas no trato digestório e reprodutor das poedeiras, possibilitando maior respaldo aos resultados obtidos.
Histologicamente, o aumento do nível de triptofano digestível levou a maior produção de albúmen pelo magno, uma vez que as glândulas tubulares do magno se apresentaram em estágio funcional mais ativo e apresentando maior quantidade de albúmen o que, permitem que a produção dos ovos seja feita em um período de tempo menor, aumentando assim a produção, peso, massa, conversão em massa e dúzia de ovos.
Ainda em relação ao magno observa-se uma maior produção de muco no epitélio desse órgão, tal característica possibilita maior rapidez na passagem do ovo pelo oviduto (Gomide-Júnior et al., 2004). Da mesma maneira, foi demonstrado que o aumento do nível de triptofano digestível também elevou a quantidade de dobras secundárias do útero das poedeiras leves em postura e gerou uma hiperplasia do epitélio desse órgão. Tais características permitiriam a formação da casca em um menor tempo, aumentando assim, a produção de ovos, visto que o ovo em formação fica no útero por tempo variável para a deposição de carbonato de cálcio (King & McLelland, 1979), sendo esta acelerada pelo aumento da superfície interna do órgão.
O melhor desempenho das aves em relação ao maior nível de triptofano nas rações também pode ser explicado pelas alterações histológicas encontradas nas vilosidades intestinais nas aves dos tratamentos que receberam rações com maiores níveis de triptofano digestível. Em tais aves, as vilosidades foram maiores e mais ramificadas, fazendo com que esse re-arranjamento da mucosa demonstre a necessidade fisiológica de ampliar a superfície absortiva do intestino delgado em relação a um determinado nutriente (Aptekmann et al., 2001). Fatos que corroboram com os resultados de Gomide-Júnior et al. (2004), nos quais demonstraram que a mucosa intestinal responde a agentes exógenos por meio de modificações morfológicas na altura e número das vilosidades intestinais, profundidade de criptas intestinais, proliferação celular e número de células mortas por perda epitelial.
Em relação ao agravamento da esteatose com o aumento dos níveis de triptofano digestível nas rações, estudos demonstram que a suplementação da ração com metionina também induz o aumento da gordura hepática (Bunchasak & Silapasorn, 2005). Este aumento da esteatose hepática pode ter sido causado pela maior síntese de estrogênio no ovário para dar suporte à maior produção de ovos, (Bunchasak & Silapasorn, 2005).
Já em relação aos pequenos depósitos de colágeno parenquimal observados no fígado das aves com o aumento dos níveis de triptofano digestível nas rações, vale ressaltar que, embora tenha sido uma pequena quantidade, este efeito denota processo fibrótico resultante de processos contínuos de regeneração hepatocitária (Fausto et al., 2003). Entretanto, como a vida produtiva de uma poedeira é pequena, tal alteração metabólica não gera perdas produtivas, qualitativas e por conseqüência, econômicas. Em adição, o tratamento que apresentou mais evidentemente essa alteração hepática, assim como maior esteatose, foi o com 0,215% de triptofano digestível, nível que não se traduziu em melhores resultados zootécnicos no experimento com relação variável. O maior acúmulo de glicogênio nos fígados (maior positividade à coloração PAS) das aves do tratamento com esse nível de triptofano digestível pode ser decorrente da presença dos depósitos de colágeno parenquimal que dificulta o escoamento do glicogênio, assim como de outros nutrientes pelas veias centrolobulares dos lóbulos hepáticos.
No experimento com relação constante, o nível de 0,214% demonstrou, na média das determinações, o melhor nível de triptofano digestível diferentemente do experimento anterior. Nos animais do segundo experimento em todos os tratamentos foram observados vilosidades intestinais compridas e ramificadas .demonstrando que o nível de 24,5% de triptofano independentemente da porcentagem constante da relação com lisina é apropriada para a maior absorção de alimentos. Foi observado também ausência de lesões hepáticas nos diferentes tratamentos, com exceção a alguns animais do tratamento com 0,165%, que apresentaram moderada esteatose, o que é considerado normal para animais de produção devido à ração hipercalórica.
Os animais dos tratamentos com 0,210 e 0,225% de triptofano, particularmente os do tratamento com 0,210% foram os que tiveram as maiores quantidades de glândulas ativas, também chamadas glândulas em regeneração, no magno, assim como, maior ramificação e comprimento das dobras uterínicas, corroborando com os índices zootécnicos que indicaram maior produção de ovos por esses animais.
Além desses resultados o aumento do triptofano mantendo constante a relação com a lisina levou a uma diminuição do peso vivo, do peso do animal sem vísceras, do fígado e principalmente da gordura visceral, demonstrando uma destinação dos nutrientes para a produção de ovos ao invés da deposição, seja em forma de músculo, gordura ou deposição de glicogênio hepático.
CONCLUSÃO
A exigência de triptofano digestível para poedeiras leves em postura é de 0,195 e 0,214% ou 205 e 219 mg/ave/dia, com relação variável e constante em 24,5%, respectivamente.
REFERÊNCIAS
AJINOMOTO. Lisina Cristal. Ajinomoto Animal Nutrition. 2010. Disponível em: htpp://www.lisina.com.br/lisina_cristal.aspx. Acessado em: 10/07/2011.
APTEKMANN, K. P.; BARALDI-ARTONI, S. M.; STEFANINI, M. A.; ORSI, M. A. Morphometric analysis of the intestine of domestic quails (Coturnix coturnix japonica) treated with different levels of dietary calcium. Anatomia, Histologia, y Embryologia, v. 30, n. 5, p. 277-280, 2001
BRASIL. Ministério da Agricultura. Equipe de pedologia e fertilidade do solo. Divisão de Agrologia - SUDENE. Levantamento exploratório: reconhecimento de solos do estado da Paraíba. Rio de Janeiro: MA/CONTA/USAID/SUDENE, 1972. 670p. (Boletim Técnico, 15).
BREGENDAHL, K.; ROBERTS, S.A.; KERR, B. et al. Ideal ratios of isoleucine, methionine, methionine plus cystine, threonine, tryptophan, and valine relative to lysine for white leghorn-type laying hens of twenty-eight to thirty-four weeks of age. Poultry Science, v.87, p.744-758, 2008.
BUNCHASAK & TAWEESAK. Silapasorn.Effects of adding methionine in low-protein diet on production performance, reproductive organs and chemical liver composition of laying hens under tropical conditions. International Journal of Poultry Science, 4 (5): 301-308, 2005.
CENTRAAL VEEVOEDERBUREAU (CVB). 1996. Aminozurenbehoefte van leghennen en vleeskuikens [Amino acid requirements for laying hens and broiler chickens]. Documentation Report nr. 18 (in Dutch), Centraal Veevoederbureau, Lelystad, the Netherlands.
COON, C., AND B. ZHANG. 1999. Ideal amino acid profile for layers examined. Feedstuffs 71(14):13–15, 31.
COSTA, F.G.P.; LIMA, M.R.; SÁ, L.M. et al. Relação triptofano digestível: lisina digestível sobre o desempenho de poedeiras leves. In: 47A REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 2010, Salvador. Anais...Salvador: SBZ, 2010.
DEPONTI, B.J.; FARIA, D.E.; FARIA FILHO, D.E. et al. Exigências de triptofano e padrão de recuperação do desempenho de poedeiras comerciais após alimentação com rações deficientes em triptofano. Revista Brasileira de Zootecnia, v.36, n.5, p. 1324-1330, 2007.
FAUSTO N. New perspectives on liver regeneration. Hepatology, 6(2):326-7, 2003.
GOMIDE-JUNIOR, M. L; STERZO, E. V.; MACARI, M.; BOLELI, I. C. Use of scanning electron microscopy for the evaluation of intestinal epithelium integrity. Revista Brasileira de Zootecnia, v. 33, n. 6, p. 1500-1505, 2004.
GONDIM, A. W. A.; FERNANDES, B. Probabilidade de chuvas para o município de Areia-PB. Agropecuária Técnica, Areia, v. 1, n.1, p. 55-63, 1980. HARMS, R.H. and RUSSEL, G.B. Evaluation of tryptophan requirement of the commercial layer by using a corn-soybean meal basal diet. 2000. Poultry Science, 79:740-742.
KING AS, MCLELLAND J. (1979) Form and Function in Birds. London: Academic Press.
LEESON, S., AND J. D. SUMMERS. 2005. Commercial poultry production. 3rd ed. University Books, Guelph, Ontario, Canada
NRC. 1994. Nutrient Requirements of Poultry. 9th ed. National Academics Press, Washington, DC.
PEGANOVA, S. & EDER, K. Studies on requirement and excess of isoleucine in laying diets. Poultry Science, 81:1714-1721, 2002.
PEGANOVA, S.; EDER, K. Interactions of various supplies of isoleucine, valine, leucine and tryptophan on the performance of laying hens. Poultry Science, v.82, p.100-105, 2003.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. Viçosa, MG: Universidade Federal de Viçosa, 2011.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L.; GOMES, P.C.; OLIVEIRA, R.F.; LOPES, D.C.; FERREIRA, A.S.; BARRETO, S.L.T. Tabelas Brasileiras para Aves e Suínos: Composição de Alimentos e Exigências Nutricionais. 2ª ed. UFV/DZO, 2005, 186p








.jpg&w=3840&q=75)